



درشت مولکول چیست؟ – به زبان ساده + تعریف و مثال
«درشت مولکولها» (Macromolecule) مولکولهای بسیار بزرگ و پیچیدهای هستند که در اثر «بسپارش» یا «پلیمریزاسیون» (Polymerization) مولکولهای کوچکتری به نام «تکپار» یا «مونومر» (Monomer) به وجود میآیند. در این مطلب به بررسی ۴ خانواده مهم از درشت مولکول های زیستی میپردازیم و در ادامه با مثالهای متنوعی درک بهتر و عمیقتری از موضوع خواهیم داشت.
- با تعریف علمی درشتمولکولها و چهار گروه اصلی آنها آشنا میشوید.
- خواهید آموخت که ساختار و نقش بیولوژیکی کربوهیدراتها و پروتئینها چگونه است.
- تفاوت ساختار، ویژگی و عملکرد پلیمرهای زیستی را با مثالهای کاربردی بررسی میکنید.
- نقش پیوندهای شیمیایی و ایزومری در ساختار و خواص مولکولها را میآموزید.
- نقش واکنشهای زیستی مانند هیدرولیز در عملکرد درشتمولکولها را تحلیل خواهید کرد.
- کاربردهای صنعتی و زیستی درشتمولکولها و روشهای جداسازی آنها را فرا میگیرید.




درشت مولکول چیست ؟
«هرمان اشتاودینگر» (Herman Staudinger) شیمیدان آلمانی و برنده جایزه نوبل سال ۱۹۵۳، مولکول را بهصورت ساختاری تعریف کرد که از بیش از یک اتم تشکیل شده باشد. به همین ترتیب درشت مولکول ها، مولکولهایی هستند که دارای بیش از ۱۰۰۰۰ اتم در ساختار خود باشند.

گاهی از درشت مولکول ها با نام «پلیمر» (Polymer) نیز یاد میشود که از بسپارش مولکولهایی حاوی عنصرهای کربن، هیدروژن و اکسیژن به وجود میآیند. واحدهای مونومری موجود در طبیعت «قطبی» (Polar) هستند و در دو سمت خود (سر و دم) دارای ویژگیهای شیمیایی و فیزیکی متفاوتی هستند.
درشت مولکول ها در زیستشناسی از اهمیت بسیاری برخوردارند. این درشت مولکول های زیستی از چهار دسته اصلی تشکیل میشوند.
- «کربوهیدارت» (Carbohydrate)
- «اسید نوکلئيک» (Nucleic Acid)
- «پروتئین» (Protein)
- «لیپید» (Lipid)
البته توجه داشته باشید که درشتمولکولها لزوما پلیمر نیستند که در فهرست بالا نیز میتوانید این نکته را در نظر بگیرید. در بخشهای بعدی هر کدام از این درشت مولکول ها را جداگانه مورد بررسی قرار میدهیم.
تکپار و بسپار
درشت مولکول ها درواقع بسپارهایی هستند که از تعداد بسیار زیادی واحد کوچکتر به نام تکپار تشکیل شدهاند. کربوهیدارتها، پروتئینها و نوکلئیک اسیدها نیز همگی بسپار هستند.

بنابراین بسپار نام دیگر درشت مولکول است. واژه Poly پیشوندی یونانی به معنای واحدهای زیاد است. یعنی هر بسپار از تعداد زیادی واحد کوچکتر از خود تشکیل شده است.

همچنین واژه Mer نیز به معنی واحد و Mono به معنی یک است. بنابراین وقتی از Polymer یا بسپار استفاده میکنیم، منظور درشت مولکولی است که از واحدهای تکرارشونده زیادی به نام Monomer یا تکپار ساخته شده است.
انواع درشت مولکول زیستی
در بخش قبلی در مورد چیستی درشت مولکول ها صحبت کردیم و انواع مولکولهای آن را معرفی کردیم. حال میخواهیم ویژگیهای هر کدام از این درشت مولکول ها را مورد بررسی قرار دهیم.
کربوهیدارت
کربوهیدارتها که به نام قند نیز شناخته میشوند، بسپارهایی از سه عنصر کربن، اکسیژن و هیدروژن هستند. نامهای عمومی کربوهیدارتها شامل قند، نشاسته، ساکارید و پلیساکارید است. عبارت ساکارید از واژه لاتین «Sacchararum»، به معنای طعم شیرینی شکر گرفته شده است.
همچنین کربوهیدارت به معنای هیدراتهایی از کربن است. این درشت مولکول ها را میتوان در سه دسته، طبقهبندی کرد.

- «مونوساکارید» (Monosaccharide)
- «دیساکارید» (Disaccharide)
- «پلیساکارید» (Polysaccharide)
این ترکیبها در نشاسته، میوه و سبزیجات، شیر و شکر یافت میشوند و منبعی برای یک تغذیه سالم هستند و از فراوانترین ترکیبهای آلی هستند که در ارگانسیمهای زنده یافت میشوند.

کربوهیدراتها از واکنش «فتوسنتز» (Photosynthesis) به وجود می آیند که واکنش «تراکم کاهشی گرمازایی» (Endothermic Reductive Condensation) روی کربن دیاکسید است. واکنش فتوسنتز نیازمند انرژی نور و رنگدانه کلروفیل است. این واکنش را میتوان بهصورت زیر نمایش داد.
درشت مولکول ها از آنجا که بسپار هستند از تعداد زیادی تکپار ساخته شدهاند بنابراین در اثر شکست درشت مولکول ها، حاصل تکپارهای زیادی است.
«متابولیسم» (Metabolism) فرایندی است که درون بدن انسان رخ میدهد و طی آن غذا تبدیل به انرژی میشود. وقتی غذا متابولیزه میشود، بدن درشت مولکول های آن را تبدیل به واحدهای کوچکتری میکند. این واحدهای کوچکتر نیروی پیشبرنده انسان طی فعالیتهای او در طول روز هستند.
کربوهیدارتها شامل یکی از معروفترین درشت مولکول های موجود، «گلوکز» (Glucose) یا «قند خون» (Blood Suger) هستند. بسیاری از کربوهیدراتها با تجزیه به اجزای کوچکتر خود، گلوکز را بهجا میگذارند.
بنابراین بدن برای متابولیز کربوهیدارتها باید زنجیرههای گلوکزی را بشکند. آنزیم «آمیلاز» (Amylase) در این راه به کمک بدن میآید. این آنزیم به زنجیرههای گلوکزی میچسبد و واحدهای گلوکزی را از یکدیگر جدا میکند. در نتیجه این فرایند انرژی مورد نیاز بدن فراهم میشود. همچنین مقداری آب و کربن دیاکسید نیز باقی میماند.
در ادامه میخواهیم با جزئیات برخی از ساختارهای قندی را مورد بررسی قرار دهیم. پیش از آن نیاز است که دو مفهوم مهم در مورد قندها را نیز مرور کنیم.
قندها با توجه به ساختار خود به دو گروه «قند کاهنده» (Reducing Sugar) و «قند غیرکاهنده» (Non-Reducing Sugar) تقسیمبندی میشوند.
- قند کاهنده: قندی که دارای گروه هیدرکسیل آزاد متصل به کربن آنومری خود باشد.
- قند غیرکاهنده: قندی که فاقد گروه هیدروکسیل آزاد روی کربن آنومری خود باشد.
برای شناسایی قندهای کاهنده از دو ترکیب «شناساگر فهلینگ» (Fehling's Reagent) و «محلول بندیکت» (Benedist's Reagent) نیز میتوان استفاده کرد بهصورتی که اگر قندی با این دو ترکیب اکسید شود، کاهنده خواهد بود. در این دو واکنش کاتیون و کاهش مییابند و رسوب نقره یا اکسید سرب از خود به جا میگذارند.
گلوکز
معروفترین کربوهیدرات گلوکز نام دارد که میتوان فرمول شیمیایی آن را بهصورت نوشت. این کربوهیدرات یک مونوساکارید «آلدوکسوزی» (Aldohexose) است، یعنی قندی ۶ کربنی آلدوزی است.
همچنین گلوکز قندی کاهنده است. همانطور که اشاره کردیم، قندهای کاهنده دارای گروه هیدروکسیلی آزادی روی کربن آنومری خود هستند.

این ماده میتواند وارد واکنش با مواد مختلفی شود و محصولات متنوعی نیز ایجاد کند. در زیر به تعدادی از این واکنشها بهصورت خلاصه اشاره شده است.
- گلوکز در واکنش با سدیم بورهیدرید سوربیتول با فرمول شیمیایی را تولید میکند. سپس سوربیتول در حضور مقادیر مازاد استیک انیدرید به فرمول و پیریدین یک هگزااستات را به وجود میآورد.
- هگزان محصول واکنش بین گلوکز و هیدروژن یدید در حضور منبع گرما است.
- گلوکز میتواند با هیدروژن سیانید وارد واکنشی برگشتپذیر شود و یک سیانوهیدرین با فرمول شیمیایی را به دست دهد.
- این ماده در حضور مقادیر مازاد استیک انیدرید به فرمول و پیریدین وارد واکنشی شود که منجر به تولید یک پنتااستات شود.
- اکسایش گلوکز در حضور در آب منجر به تولید گلوکونیک اسید با فرمول شیمیایی میشود.
- اکسایش این ماده در حضور نیتریک اسید رقیق محصول گلوکارید اسید را به دست میدهد.
از هیدروژن یدید گرم معمولا برای حذف کاهشی گروههای عاملی اکسیژن در مولکولها استفاده میشود اما در مورد گلوکز، نتیجه این واکنش محصول هگزان با بازده بسیار پایین بود. از این مشاهده اینطور برداشت شد که ۶ کربن این ساختار در زنجیرهای خطی و غیرشاخهای حضور دارند. وجود گروه عاملی کربونیلی آلدهیدی از تشکیل سیانوهیدرین، کاهش گلوکز به هگزاالکل سوربیتول و اکسایش ملایم با محصول گلوکونیک اسید، استنتاج شد. همچنین اکسایش قوی با نیتریک اسید رقیق که منجر به تولید محصول گلوکاریک اسید میشود نیز وجود زنجیره ۶ کربنی را تایید میکند.
گلوکز و ساکاریدهای دیگر به دلیل داشتن استخلافهای «دیاُل» (Diol) مجاور هم با استفاده از «پریودیک اسید» (Peroiodic Acid) شکسته میشوند. این واکنش که به نام «واکنش مالاپراد» (Malaprade Reaction) شناخته میشود برای آنالیز انتخابی مشتقهای دارای استخلاف اکسیژنی ساکاریدها بسیار مفید و کاربردی است. واکنش مالاپراد را میتوان بهصورت زیر نمایش داد.
پیکربندی گلوکز
از آنجا که این مولکول دارای ۴ «مرکز کایرال» (Chiral Center) است، میتواند ۱۶ ایزومر فضایی متنوع داشته باشد یعنی ۸ جفت «دیاسترومری» (Diastereomer) یا ۱۶ «انانتیومر» (Enantiomer) وجود خواهد داشت. اگر مولکولی داری مرکز کایرال باشد، برای محاسبه تعداد ایزومرهای ممکن آن میتوان از رابطه زیر بهره برد.
سوالی که به وجود میآید این است که از بین این ۱۶ ایزومر با فرمول شیمیایی یکسان کدام یک به گلوکز تعلق دارد. این سوال را شیمیدان آلمانی «امیل فیشر» (Emil Fischer) در سال ۱۸۹۱ پاسخ داد. او نحوه نوینی برای نشان دادن آسان ایزومرها به وجود آورد و در سال ۱۹۰۲ به همین خاطر برنده جایزه نوبل شیمی شد. این نحوه نمایش را امروزه به نام فرم فیشر میشناسیم.
زمانی که فیشر روی پیکربندی گلوکز تحقیقات میکرد، تعیین پیکربندی مطلق یک انانیتومر ممکن نبود. به همین دلیل فیشر روش جدیدی برای نامگذاری انتخاب کرد. او به +-گلوکز و آلدوزهای دارای پیکربندی مشابه واژه را نسبت داد و آنها را در یک خانواده دستهبندی کرد. به تصویر آینهای این ایزومرها نیز واژه را به عنوان پیشوند نسبت داد. این ایزومرها را میتوانید در تصویر زیر مشاهده کنید. در تمام ساختارها کربنهایی که کایرال هستند با رنگ قرمز متمایز شدهاند. آخرین مرکز کایرال در یک زنجیره آلدوزی (دورترین از گروه آلدهیدی) مبنایی برای نامگذاری ایزومرها با پیشوند و است. اگر دورترین گروه هیدروکسیلی از گروه آلدهید در فرم فیشر به سمت راست جهتگیری کرده باشد، عضوی از خانواده خواهد بود و بالعکس، یعنی اگر دورترین گروه هیدروکسیلی از گروه آلدهید در فرم فیشر به سمت چپ جهتگیری کرده باشد، آن ایزومر عضوی از خانواده است.

امروزه نیز از همین سیستم استفاده میشود زیرا در سال ۱۹۵۱ دانشمندی هلندی با مطالعه به روش فلورسانس اشعه ایکس تارتاریک اسید، درستی روش فیشر را تایید کرد. به این نکته بسیار مهم توجه داشته باشید که بین پیشوند و در نامگذاری مولکولها و جهت چرخش ویژه نوری آنها رابطهای وجود ندارد. چرخش نوری ویژه را میتوان با پلاریمتر تعیین کرد اما برای تعیین پیکربندی مطلق معمولا نیاز به واکنشهای شیمیایی داریم که فضاویژه باشند.
فرم های آنومری گلوکز
یافتههای علمی معمولا توسط یک شخص و در یک زمان تکمیل نمیشوند، بلکه مرحله به مرحله و با مشارکت دانشمندان و تحقیقات متعددی به انجام میرسند. گلوکز نمونهای از این موارد است که اطلاعات پیرامون آن در طی زمان و توسط افراد مختلف یافته و تکمیل شده است.
مطالعات فیشر روی ساختار فضایی گلوکز نتوانست تمامی ابهامات را پیرامون آن از بین ببرد و در سال ۱۸۹۵ گزارش شد که دو بلور متفاوت گلوکز به دست آمدهاند. هر دوی این بلورها واکنشهای مورد انتظار از گلوکز را انجام میدادند و با نسبتهای برابری در آب حل میشدند. زمانی که گلوکز تبدیل به پنتامتیل اتر همتای خود شد، دو ایزومر متفاوت به دست آمد که هیچ کدام واکنشهای آلدهیدها را انجام نمیدادند.
به تصویر زیر دقت کنید. در آن چرخش پیرامون کربن شماره ۴ و کربن شماره ۵ باعث نزدیک شدن گروه عاملی هیدروکسیل روی کربن شماره ۵ به کربن آلدهیدی میشود و برای راحتی همیاستال تشکیل شده تخت نشان داده شده است اما درواقع دارای «صورتبندی صندلی» (Chair Conformatioin) است. اتم کربن همیاستالی شماره ۱ تبدیل به مرکز فضایی جدیدی میشود که از آن با عنوان کربن آنومری نام برده میشود. توجه داشته باشید که گروه روی کربن آنومری میتواند در دو جهت بالا و پایین جهتگیری داشته باشد و دو آنومر متفاوت را تشکیل بدهد. ابن دو حالت متفاوت، با نامهای آلفا و بتا از یکدیگر مجزا میشوند.

حال میتوان به پاسخ ابهام بالا نزدیک شد. میدانیم که همیاستالها در محلول با فرم الکلی و کربونیلی خود در تعادل هستند. متعاقبا محلول آلفا-گلوکز و بتا-گلوکز در محلول تعادلی هر دو فرم آنومری نشان داده شده در تصویر بالا را به وجود میآورند و حالت خطی را نیز در عین حال خواهند داشت. توجه داشته باشید که با وجود غلظت بسیار پایین آلدهید خطی در ترکیب، همچنان واکنشهای شیمیایی آلدهید را انجام میدهد.
واکنش های مهم کربوهیدرات
امیل فیشر روی بسیاری از واکنشهای کربوهیدارتها نیز مطالعه انجام داده است. در این بخش میخواهیم برخی از مهمترین این واکنشها را مورد بررسی قرار دهیم.
اکسایش
زمانی که گروه عاملی آلدهید یک قند آلدوز اکسید میشود، محصول آن که با نام «آلدونیک اسید» (Aldonic Acid) شناخته میشود، دارای کربوکسیلیک اسید است. از آنجا که الکل نوع دو نیز در این ترکیبها وجود دارد، میتوان از عوامل اکسیدکننده ملایمی مانند هیپوبرومیت نیز استفاده کرد. اگر هر دو سر آلدوز اکسید شود و کربوکسیلیک اسید تولید کند، محصول «آلدریک اسید» (Aldaric Acid) خواهد بود. با تبدیل آلدوز به آلدریک اسیدش، پایان زنجیره در دو طرف مولکول با یکدیگر برابر خواهد بود و این میتواند باعث ایجاد تقارن در تمام نقاط این ساختار شود. بنابراین ریبوز، زایلوز، آلوز و گالاکتوز، آلدریک اسیدی غیرکایرال را به دست میدهند و همانطور که میدانید مولکولهای غیرکایرال از نظر نوری فعال نیستند. برای روشن شدن موضوع به واکنشهای تصویرهای زیر دقت کنید.

برخی قندهای آلدوزی میتوانند آلدریک اسید کایرال مشابهی به وجود بیاورند. مثالی از این مورد دو قند آرابینوز و لایزوز هستند. این مسئله را میتوانید در تصویر زیر مشاهده کنید.

کاهش
کاهش قند آلدوز به کمک سدیم بورهیدرید منجر به تولید یک «آلدیتول» (Alditol) میشود بنابراین مانند اکسایش شامل تغییر در پیکربندی میشود. در جدول زیر محصولات واکنشهای کاهش و اکسایش را مشاهده میکنید.
| فرایند | محصول | فرمول شیمیایی |
| کاهش با | آلدیتول | |
| اکسایش با | آلدونیک اسید | |
| اکسایش با | آلداریک اسید |
انواع کربوهیدارت
کربوهیدارتها را با توجه به ویژگیهایشان به دستههای متنوعی تقسیمبندی میکنند. یکی از این دستهبندیها قندها را با توجه به تعداد تکپارهایشان میسنجد. در این مورد میتوان به گزینههای زیر اشاره کرد.
- مونوساکارید: این قندها تنها از یک تکپار قندی تشکیل شدهاند.
- دیساکارید: به موادی گفته میشود که شامل دو تکپار قندی باشند.
- پلیساکارید: به تمام قندهایی که دارای بیش از دو تکپار قندی باشند، گفته میشود.
کتوز
اگر مونوساکاریدی دارای گروه کربونیلی در یکی از اتمهای کربن میانی زنجیره خود باشد، به آن کتوز میگویند. این گروه کربونیلی معمولا روی کربن شماره ۲ دیده میشود. برای درک بهتر تصاویری در ادامه خواهیم آورد که در آن کربنهای کایرال با رنگ قرمز مشخص شدهاند. همانطور که انتظار میرود، گروه کربونیلی کتوز میتواند با سدیم بورهیدرید کاهش پیدا کند و محصول این واکنش معمولا مخلوطی از ترکیبات اپیمری است. دی-فروکتوز که شیرینترین قند طبیعی است را در نظر بگیرید. این قند کاهش پیدا میکند و مخلوطی از دی-گلوسیتول (سوربیتول) و دی-مانیتول را بر جا میگذارد. یکی از محصولات این واکنش یعنی مانتیول خود کربوهیدارتی طبیعی است.

با وجود اینکه کتوزها و آلدوزها مونوساکاریدهای متفاوتی هستند، ویژگیهای شیمیایی آنها به دلیل سهولت تبدیل آنها به یکدیگر در حضور کاتالیزور بازی و اسیدی، شباهتهای فراوانی دارند. این ترتیب از طریق توتومریزاسیون ان-دیاُل صورت میگیرد.
همانطور که مشاهده میکنید اکسیژن حلقه در پشت صفحه و دور از بیننده قرار دارد. کربن آنومری که با رنگ قرمز نشان داده شده است، در سمت راست قرار دارد. توجه داشته باشید که محصول به دو فرم آنومری بتا و آلفا حضور دارد. در فرم آلفا گروه عاملی به سمت بالا و در فرم بتا به سمت پایین قرار دارد. به تفاوت این دو فرم در تصویر خوب دقت کنید.
فرم های حلقوی مونوساکارید
فرم مطلوب بسیاری از مونوساکاریدها، همیاستال حلقوی آنها است. حلقههای ۵ و ۶ عضوی نسبت به باقی حلقههای احتمال حضور بیشتری دارند زیرا در حلقههای کوچکتر وجود فشار زاویهای بالا، تشکیل آن را نامطلوب میکند. به حلقههای ۵ عضوی «فورانوز» (Furanose) و به حلقههای ۶ عضوی «پیرانوز» (Pyranose) میگویند. ریبوز که عضو مهمی از آلدوپنتوزها است، معمولا ساختاری فورانوزی اتخاذ میکند. این تبدیل را میتوانید در تصویر زیر مشاهده کنید.

پیرانوزهای حلقوی بسیاری از مونوساکاریدها را بهصورت نشان داده شده در تصویر بالا نمایش میدهند. به این فرم «طرح هاوارث» (Haworth Formula) گفته میشود که به نام شیمیدان بریتانیایی «نورمن هاوارث» (Norman Haworth) نامگذاری شده است. مانند حلقه فورانوزی تصویر قبل، کربن آنومری در سمت راست و اکسیژن حلقه در قسمت پشت و دور از بیننده قرار دارد. در خانواده قندهای اتصال بهصورت بتا و آلفا مانند حلقه فورانوزی خواهد بود. (جهتگیری بتا رو به بالا و جهتگیری آلفا رو به پایین است.)
توجه داشته باشید که این طرح می تواند رابطه بین دو فرم آنومری را بسیار خوب نشان دهد اما شکل واقعی مولکول این نیست زیرا این مولکولها درواقع به فرم پایداری صندلی خود حضور دارند. در تصویر زیر میتوانید طرح هاوارث ۴ پیرانوز را با فرم ایزومر صندلی آن مقایسه کنید. کربنهای آنومری موجود در ساختارها با رنگ سرخابی مشخص شدهاند.

اندازه همیاستالهای حلقوی که یک قند میتواند داشته باشد ثابت نیست بلکه با تغییر استخلافها و ویژگیهای ساختاری میتواند تغییر کند. آلدوهگزوزها معمولا حلقه پیرانوزی تشکیل میدهند و برای همتای پنتوزی آنها تشکیل پنتوز به فورانوز ارجح است اما در این موارد میتوان استثناهایی نیز یافت.
برای دانستن اندازه حلقه بسیاری از مونوساکاریدها میتوان از شکست اکسایشی مشتقات پرمتیله استفاده کرد. همانطور که گفتیم به دلیل وجود فشار زاویهای، تشکیل حلقههای ۵ و ۶ عضوی ارجح هستند. در تصویر زیر توالی واکنشهایی که میتوان برای پی بردن به اندازه گلوکز از آنها استفاده کرد، آورده شده است.

یکی از گروههای متیل اتر در این مشتق بخشی از استال است و بنابراین بهسرعت در محلول اسیدی، هیدرولیز میشود. سپس فرم خطی این تترامتیل گلوکز به کمک نیتریک اسید به حد واسطی کتو-اسیدی تبدیل میشود. موقعیت کربونیل کتونی نشاندهنده اندازه هتروسیکل اولیه است که برای حلقه پیرانوزی کربن شماره ۴ و برای حلقه فورانوزی کربن شماره ۳ است. اکسایش بعدی ساختار را از جایی میشکند که منجر به تولید گروه کربونیلی شود. اکسایش انتهایی مخلوطی از دو کربوکسیلیک اسید را میدهد که زنجیرهشان به اندازه یک کربن با یکدیگر متفاوت هستند. زمانی که این قطعات شناسایی شوند، موقعیت گروه عاملی کتون مشخص میشود.
گلیکوزید
مشتقهای استالی که از واکنش بین یک الکل و یک مونوساکارید در حضور کاتالیزور اسیدی به وجود میآیند، «گلیکوزید» (Glycodie) نامیده میشوند. در نامگذاری گلیکوزیدها پسوند «اوز» (Ose) با «اوزید» (Oside) جایگزین میشود و ابتدا نام الکل آورده میشود. در واکنش تشکیل گلیکوزید یک مولکول آب آزاد میشود. محصول دیاتری ایجادشده در برابر باز و اکسیدانهای بازی مقاوم است و از آنجا که آلدوله شدن با کاتالیزور اسیدی واکنشی «برگشتپذیر» (Reversible) است، محصول گلیکوزیدی میتواند دوباره تبدیل به قند و الکل اولیه خود شود. در تصویر زیر میتوانید مراحل این فرایند را بهخوبی مشاهده کنید.

آنومرهای متیل گلیکوزید در واکنش بالا بهصورت ۶۶٪ آلفا و ۳۴٪ بتا تولید میشوند. در بخشهای قبلی گفتیم که حلقههای پیرانوزی فرم صندلی را ترجیح میدهند که در آن بیشتر استخلافها بهصورت استوایی قرار دارند. در محصول بتا تمامی استخلافها به فرم استوایی حضور دارند در حالی که استخلاف روی کربن شماره ۱ در آنومر بتا بهصورت محوری قرار گرفته است. بنابراین از آنجا که حلقه سیکلوهگزان موقعیت استوایی را به موقعیت محوری ترجیح میدهد، محصول مطلوب واکنش بتا است و این انتخابگری را به مفهوم با نام «اثر آنومری» (Anomeric Effect) نسبت میدهند.
پیوندهای گلیکوزیدی را بهوفور میتوان در سیستمهای زیستی پیدا کرد، قندهای مختلف میتوانند به درشت مولکولی مانند لیپید متصل شوند و ویژگیهای شیمیایی و انحلالپذیری آنها نیز دستخوش تغییر خواهد شد. به دلیل همین تاثیر، آنزیمهای مختلفی وجود دارند که توانایی حذف ساختارهای قندی از الکل، فنول و آمینها را دارند. این آنزیمها با نام «گلیکوزیداز» (Glycosidase) شناخته میشوند. شیمیدانها به قسمت قندی گلیکوزیدهای طبیعی «گلیکون» (Glycon) و به قسمت الکلی آن «آگلیکون» (Aglycon) میگویند.
دی ساکارید
دیساکارید ترکیبی حاوی دو تکپار مونوساکاریدی است. به عبارتی اگر قسمت الکلی موجود در گلیکوزید توسط مونوساکارید دومی اشغال شود، نتیجه، تشکیل یک دیساکارید خواهد بود. مثلا در تصویر زیر میتوانید دیساکاریدی متشکل از دو مونوساکارید گلوکز را ببینید. در این تصویر حلقههای گلوکوپیرانوزی با دو عبارت و مشخص شدهاند و دور پیوند گلیکوزیدی نیز دایرهای آبی وجود دارد. توجه داشته باشید که این پیوند آلفا میتواند مثل مالتوز و ترهالوز از نوع آلفا باشد یا مثل سلوبیوز و ژنتیوبیوز از نوع بتا باشد. هیدرولیز این دیساکاریدها به کمک کاتالیزور اسیدی تنها یک محصول به جا میگذارد که گلوکز است. هیدرولیزهایی که به کمک آنزیم انجام میشوند نسبت به نوع پیوند گلیکوزیدی انتخابگر هستند، بهطوری که آلفا گلیکوزیداز مالتوز و ترهالوز را میشکند و گلوکز تولید میکند اما روی سلوبیوز و ژنتیوبیوز تاثیری ندارد. عملکرد آنزیم بتا گلیکوزیداز دقیقا برعکس است، یعنی روی مالتوز و ترهالوز بیاثر است و سلوبیوز و ژنتیوبیوز را تبدیل به واحد سازنده آنها یعنی گلوکز میکند.

برای رسم ساختار سلوبیوز، نیاز است که یکی از حلقههای گلوکوپیرانوز ۱۸۰ درجه چرخانده شود که معمولا برای داشتن زاویه دید بهتر این کار انجام نمیشود. پیوند بین حلقههای گلوکوپیرانوز در سلوبیوز و مالتوز از کربن آنومری حلقه به گروه هیدروکسیلی کربن شماره ۴ در حلقه است که این باعث میشود کربن آنومری حلقه دست نخورده باقی بماند. این مطالب را میتوانید در تصویر زیر مشاهده کنید.

ژنتیوبیوز دارای پیوند گلیکوزیدی بتا است که از کربن شماره ۱ حلقه به کربن شماره ۶ حلقه برقرار میشود. از آنجا که سلوبیوز، مالتوز و ژنتیوبیوز همیاستال هستند، هر سه جزو قندهای کاهنده دستهبندی میشوند. ترهالوژ قندی است که در برخی قارچها یافت میشود و یک «بیس استال» (Bis-Acetal) است. به همین دلیل نیز متعلق به خانواده قندهای غیرکاهنده است. سیستم نامگذاری قندها توانایی این را دارد که آنها را از هم بهخوبی متمایز کند اما در بسیاری مواقع به دلیل طولانی بودن اسمها در این سیستم، از نامهای رایج قندها استفاده میشود. برای نمونه در زیر نام همین ۴ قند مورد بحث را در این سیستم نامگذاری خواهیم آورد.
- سلوبیوز: ۴-او-بتا-دی-گلوکوپیرانوزیل-دی-گلوکز
- مالتوز: ۴-او-آلفا-دی-گلوکوپیرانوزیل-دی-گلوکز
- ژنتیوبیوز: ۶-او-بتا-دی-گلوکوپیرانوزیل-دی-گلوکز
- ترهالوز: آلفا-دی-گلوکوپیرانوزیل-آلفا-دی-گلوکوپیرانوزید
گرچه هر ۴ دیساکارید بالا از واحدهای گلوکوپیرانوزی تشکیل شدهاند، اما دارای تفاوتهای جالبی هستند. مالتوز که گاهی با عنوان قند مالت نیز از آن یاد میشود، در بدن انسان هضم بسیار آسانی دارد و توسط مخمر تخمیر میشود. سلوبیوز از هیدرولیز سلولز به دست میآید. این ماده طعم خاصی ندارد و بدن انسان نیر نمیتواند آن را هضم کند. همچنین سلوبیوز توسط مخمر نیز غیرقابل تخمیر است. برخی از باکتریها دارای آنزیمی به نام بتا-گلیکوزیداز هستند و توانایی هیدرولیز آن و به وجود آوردن سلولز را دارند. وجود این آنزیم در بدن گاوها و موریانهها اجازه میدهد تا از این ماده به عنوان منبعی برای تغذیه خود استفاده کنند. نکته پایانی در مورد این قندها این است که ترهالوز طعم شیرینی دارد اما ژنتیوبیوز اینطور نیست. این قند حتی طعمی تلخ دارد.

دیساکاریدهای معروف دیگری نیز وجود دارند، برای مثال لاکتوز که به آن قند شیر نیز گفته میشود. این قند از دو واحد مونوساکاریدی گالاکتوز و گلوکز تشکیل شده است که به وسیله یک پیوند گلیکوزیدی از نوع بتا به یکدیگر متصل هستند. بسیاری از بزرگسالان نمیتوانند شیر مصرف کنند زیرا دچار عدم تحمل لاکتوز هستند. این افراد آنزیم مورد نیاز برای شکستن این قند یعنی «لاکتاز» (Lactase) را در بدن خود ندارند یا به مقدار بسیار کمی دارند. به همین دلیل بعد از مصرف شیر علائمی مانند اسهال و دلدرد را تجربه میکنند. باقی لبنیات مانند پنیر مقدار لاکتوز بسیار کمتری دارند و برای این افراد مشکلی ایجاد نمیکنند.

ساکاروز که از نیشکر و چغندر به دست میآید، پرمصرفترین قند است که به عنوان شیرینکننده مورد استفاده قرار میگیرد. این قند غیرکاهنده است و از دو واحد مونوساکاریدی گلوکز و فروکتوز تشکیل شده است. پیوند گلیکوزیدی این قند دو کربن آنومری مونوساکاریدها را به یکدیگر متصل میکند.

پلی ساکارید
همانطور که از نام این ترکیب پیداست، پلیساکارید درشت مولکولی با جرم مولکولی بالا است که از تعداد زیادی واحدهای مونوساکاریدی به وجود آمده است. گاهی از نام «گلیکان» (Glycan) نیز برای این ترکیبها استفاده میشود. معروفترین درشت مولکول های پلیساکاریدی شامل سلولز، نشاسته و گلیکوژن هستند که همگی از گلوکز تشکیل شدهاند و این امر با هیدرولیز این ترکیبها بهراحتی مشاهده میشود. از آنجا که هیدرولیز جزئی سلولز مقادیر متفاوتی از سلوبیوز را به دست میدهد، میتوان اینطور برداشت کرد که واحدهای گلوکزی در این درشت مولکول توسط پیوند گلیکوزیدی بتا بین کربن شماره ۱ و کربن شماره ۴ قند مجاور تشکیل شده است. درشت مولکولهایی متشکل از مونوساکاریدهایی مانند مانوز، گالاکتوز، زیلوز و آرابینوز نیز وجود دارند. در تصویر زیر ساختار یکی از این درشت مولکول های پلیساکاریدی به نام سلولز آورده شده است.

سلولز بهوفور در کره زمین و در مواد مختلف مشاهده میشود. الیاف طبیعی پنبه سلولز خالص هستند و چوب بوتهها و درختان حدود ۵۰٪ سلولز دارند. بسته به منبعی که سلولز در آن موجود است، این بسپار گلوکزی با فرمول کلی دارای بین ۵۰۰ تا ۵۰۰۰ است. همانطور که در تصویر بالا مشاهده میکنید، واحدهای مونوساکاریدی بهصورت خطی به یکدیگر متصل شدهاند. وجود پیوند گلیکوزیدی بین گلوکزها اجازه کش آمدن را به این زنجیره میدهد. از طرفی این ساختار به کمک پیوندهای هیدروژنی درونمولکولی به پایداری میرسد.
اگرچه یک پیوند هیدروژنی تنها پیوند قوی نیست اما وجود تعداد زیادی از آن به پایداری درشت مولکول ها کمک قابلتوجهی میکند. بسیاری از حیوانات توانایی هضم این درشت مولکول قندی را ندارند و انسان نیز بیشتر سلولز موجود در مواد غذایی خود را بدون تغییر دفع میکند.

سلولز معمولا با بسپار دیگری به نام «همیسلولز» (Hemicellulose) همراه است که ساختاری سبکتر و شاخهای دارد. برخلاف سلولز، همیسلولز ساختار ضعیفی دارد و بهراحتی توسط اسید و باز ضعیف هیدرولیز میشود. همیاستال از قند دی-پنتوز و زایلوز تشکیل شده است.

نشاسته
«نشاسته» (Starch) درشت مولکولی قندی است که در ریشه، ساقه، غده و پیاز گیاهان یافت میشود. بیشتر حیوانات از جمله انسانها به این ماده مغذی در رژیم غذایی خود وابستگی دارند. ساختار نشاسته از سلولز پیچیدهتر است و در آب سرد حل نمیشود. نشاسته از دو ماده اصلی تشکیل شده است.
ماده اول که در آب انحلالپذیری خوبی دارد را با نام «آمیلوز» (Amylose) میشناسیم. مولکول آمیلوز ساختاری خطی دارد که از هزاران واحد گلوکزی تشکیل شده است. در این ساختار پیوندهای بین گلوکز از نوع آلفا و از نوع کربن ۱-کربن ۴ است. ماده دوم که جرم مولکولی بسیار بالاتری دارد و از حدود یک میلیون واحد گلوکزی تشکیل میشود، «آمیلوپکتین» (Amylopectin) نامیده میشود. این ماده شبکهای شاخهدار است که از پیوندهای گلیکوزیدی کربن ۱-کربن ۴ و کربن ۱-کربن ۶ تشکیل شده است که در آب حل نمیشود.

هیدرولیز نشاسته معمولا با واکنشی آنزیمی انجام میشود و منجر به تولید شیرهای میشود که میزان بسیار زیادی گلوکز دارد. زمانی که ماده اولیه این واکنش نشاسته ذرت باشد، محصول نهایی شیره ذرت خواهد بود. شیره ذرت به میزان زیادی در نرم کردن بافتها، افزایش حجم، جلوگیری از تشکیل بلور و تقویت طعم مواد خوراکی مورد استفاده قرار میگیرد.

گلیکوژن منبعی برای ذخیره گلوکز در بدن حیوانات است. ساختار گلیکوژن مشابه آمیلوپکتین است فقط در شاخهها با یکدیگر تفاوتهایی دارند.
سنتز ترکیبات سلولزی
همانطور که پیشتر اشاره کردیم، الیاف طبیعی نخی که پرکاربردترین پارچه است، تقریبا سلولز خالص است. تولید این پارچه شامل چند مرحله فیزیکی است.
- پنبهزنی
- شانه زدن
- نخریسی
بهترین پارچه نخی از الیافی به دست میآید که دارای رشتههای بلند باشند و گردوغبار آنها گرفته شده باشد. از سلولز خام موجود در چوب که کیفیت پایینتری دارد، برای تولید کاغذ استفاده میشود. برای اینکه قادر باشیم از سلولز به طرق مختلف و کارامدتری استفاده کنیم، چند راه حل شیمیایی جلوی پا داریم. در تمامی این تبدیلها سه گروه هیدروکسیلی آزاد روی گلوکزهای زنجیره سلولزی، نقشی اساسی بازی میکنند. «استریفیکاسیون» (Sterification) آنها منجر به تولید محصولات بسپاری میشود که ویژگیهای بسیار متفاوتی با درشت مولکول سلولز دارند.

سلولز نیترات
سلولز نیترات اولین بار حدود ۱۵۰ سال پیش از واکنش بین نیتریک اسید و سلولز به وجود آمد. این ماده اگر تماما نیتریله شود، تبدیل به میشود که «پنبه باروتی» (Guncotton) نامیده میشود و بسیار اشتعالپذیر است. این ماده در تولید «پودر بیدود» (Smokeless Powder) مورد استفاده قرار میگیرد. اگر سلولز بهصورت جزئی نیتریله شود، نتیجه محصولی خواهد بود که آن را «پیروکسیلین» (Pyroxylin) مینامیم. این ماده در اتر حل میشود و در بازهای از زمان در فیلم عکاسی و لاک مورد استفاده قرار میگرفت اما اشتعالپذیری بالا، مصرف آن را محدود کرد.
بهعلاوه این ماده در طول زمان، هیدرولیز و تبدیل به نیتریک اسید میشود. این باعث کاهش کیفیت تصاویر مورد استفاده در فیلمها میشد.

سلولز استات
این ماده دارای واحدهای تکرارشونده است و نسبت به پیروکسیلین اشتعالپذیری بسیار کمتری دارد. به همین دلیل جایگزین آن در بسیاری مصارف شده است. این درشت مولکول از واکنش سلولز و استیک انیدرید تحت کاتالیزوری اسیدی به دست میآید. ویژگیهای محصول این واکنش با توجه به درجه «استیله کردن» (Acetylation) تغییر میکند.

ایزومر کربوهیدارت
در این بخش میخواهیم بهصورت خلاصه برخی از ایزومرهای ساختارهای قندی را مورد بررسی قرار دهیم. گلیسرآلدهید میتواند به دو فرم ایزومری وجود داشته باشد که این دو تصویر آینهای یکدیگر هستند. این دو فرم را میتوانید در تصویر زیر مشاهده کنید. در حالتی که گروه آلدهیدی رو به بالا باشد، گروه باید در سمت چپ مولکول باشد تا پیکربندی ایزومر از نوع باشد. شیمیدانها این مولکول را مبنایی برای تشخیص ایزومرهای فضایی باقی مولکولهای کربوهیداتی در نظر گرفتهاند. تمام مونوساکاریدهای طبیعی متعلق به خانواده ایزومر نوری هستند. به این نکته توجه داشته باشید که به دلیل تفاوتی که آنزیمها در رفتار با ایزومرهای نوری متفاوت دارند، میتوان از آنها نیز برای تشخیص این ایزومرها بهره برد.

بنابراین مونوساکاریدهایی که گروه متصل به کربن کایرال آنها که دورترین از گروه کربونیلی است در سمت راست قرار داشته باشد، به خانواده تعلق دارند. به دلیل عدم توانایی فرم نمایشی گلوله و میله، برای درک بهتر ساختار این ایزومرها از فرم فیشر استفاده میشود.

در این بخش میخواهیم منشا و دلیل نامگذاری برخی از معروفترین قندها را بیان کنیم.
مونوساکاریدها
- گلوکز: از کلمه یونانی به معنای نوشیدنی شیرین گرفته شده است. این قند مسئول طعم شیرین انگور است و مادهای است که در بدن از آن به عنوان قند خون یاد میشود.
- گالاکتوز: از کلمه یونانی Galat به معنای شیر گرفته شده است. این ماده مونوساکارید سازنده قند شیر، لاکتوز است.
- فروکتوز: این قند از کلمه Fructus که در یونانی معنای میوه میدهد، گرفته شده است. فروکتوز در میوه و عسل یافت میشود و شیرینترین طعم را در میان قندها دارد.
- ریبوز: این قند در اسکلت ساختار دو مولکول مهم زیستی RNA و DNA وجود دارد.

دیساکاریدها
در این بخش به معرفی چند دیساکارید میپردازیم.
- ساکاروز: نام این قند از واژه فرانسوی (Sucre) که به معنای شکر است، گرفته شده است. ساکاروز دیساکاریدی متشکل از دو مونوساکارید فروکتوز و گلوکز است. ساکاروز به قند سفره معروف است.
- لاکتوز: این قند از روی واژه (Lact) که در لاتین به معنای شیر است، نامگذاری شده است. این دیساکارید موجود در شیر از گلوکز و گالاکتوز تشکیل شده است.
- مالتوز: کلمه (Malt) در زبان فرانسوی، دلیل نامگذاری این قند است. این دیساکارید از دو واحد مونوساکاریدی گلوکز تشکیل شده است. مالتوز در جوانه دانهها یافت میشود.

پلیساکارید
- نشاسته: گیاهان واحدهای مونوساکاریدی گلوکز را بهصورت نشاسته در خود ذخیره میکنند. غلاتهایی مانند برنج، گندم، ذرت، جودوسر و غدههایی مانند سیبزمینی منابعی غنی از نشاسته هستند.
- سلولز: سلولز ماده اصلی تشکیلدهنده دیواره صلب سلولهای گیاهان است و بهصورت پلیساکاریدی خطی حضور دارد. این قند نیز از واحدهای مونوساکاریدی گلوکز تشکیل شده است.
- گلیکوژن: این درشت مولکول نقش نشاسته در بافت گیاهان را برای انسانها بازی میکند و محلی برای ذخیره گلوکز است. گلیکوژن بهطور عمده در کبد و ماهیچهها سنتز میشود.

اسید نوکلئیک
اسیدهای نوکلئیک شامل «دیانای» (DNA) و «آرانای» (RNA) و بسپارهای «نوکلئوتیدی» (Nucleotide) هستند. این درشت مولکول ها ساختاری حاوی یک گروه پنتوزی (قند ۵ کربنه)، یک گروه فسفاتی و یک باز نیتروژنی میباشند.
DNA ساختاری است که تمام ویژگیهای ارثی انسان را در خود حفظ و منتقل میکند.

بازهای نیتروژنی
به دو جفت ساختار زیر که مربوط به بازهای نیتروژنی هستند، خوب دقت کنید. به دو باز نیتروژنی تکحلقه، «پیریمیدین» (Pyrimidine) و به دو باز نیتروژنی دوحلقهای، «پورین» (Purine) گفته میشود.
هر دو اینها دارای حداقل یک جایگاه هستند که میتواند به ساختاری آلی متصل شود. هر ۴ ساختار نشانداده شده «چندعاملی» (Polyfunctional) هستند و میتوانند به فرمهای «توتومری» (Tautomeric) حضور داشته باشند.

ساختار شیمیایی DNA
از نظر شیمیایی، «دیانای» (Deoxyribonucleic Acid) از یک قند پنتوزی، یک فسفریک اسید و چند باز حلقوی دارای نیتروژن ساخته شده است. قند موجود در این ساختار بهصورت «بتا-دی-۲-دئوکسی ریبوز» (β-D-2-deoxyribose) است. همچین بازهای دارای نیتروژن موجود شامل موارد زیر هستند.
- آدنین
- گوانین
- سیتوزین
- تیمین
این بازها و نحوه قرارگیری آنها نقش بسیار مهمی را در ذخیره و انتقال اطلاعات از نسلی به نسل دیگر ایفا میکند. DNA، درشت مولکولی دارای دو رشته است که این دو تکمیلکننده یکدیگر هستند.
ساختار شیمیایی RNA
این درشت مولکول نیز مانند DNA، از قند پنتوزی، فسفریک اسید و بازهای حلقوی دارای نیتروژن تشکیل میشود. قند RNA «بتا-دی-ریبوز» (β-D-ribose) است. بازهای هتروسیکلی موجود در این درشت مولکول شامل موارد زیر هستند.

- آدنین
- گوانین
- سیتوزین
- اوراسیل
همانطور که مشاهده میکنید ساختار دو درشت مولکول RNA و DNA در بازهای نیتروژنی با یکدیگر متفاوت هستند. ساختار RNA تنها دارای یک رشته است و گاهی دچار تاخوردگی میشود که منجر به تولید مارپیچ خواهد شد. RNAها به سه دسته زیر تقسیمبندی میشوند و هرکدام کارایی منحصربهفردی دارند.
- ام-آرانای (m-RNA): در اینجا m مخفف Messenger است و به این نوع «RNA پیامرسان» گفته میشود.
- آر-آرانای (r-RNA): در اینجا r مخفف Ribosomal است و به آن «RNA ریبوزومی» میگوییم.
- تی-آرانای (t-RNA): t مخفف Transfer است و به این نوع «RNA ناقل» گفته میشود.
در شکل زیر ساختار این دو درشت مولکول را مشاهده میکنید. به تفاوت موجود میان بازهای نیتروژنی هرکدام دقت کنید.

عملکرد نوکلئیک اسیدها
این درشت مولکول ها نقش بسیار پررنگ و مهمی را در بدن انسان ایفا میکنند، بهطوری که بدون آنها و عملکردشان، ادامه حیات ممکن نخواهد بود. در ادامه به برخی از آنها بهصورت خلاصه اشاره میکنیم.
- این درشت مولکول ها در انتقال ویژگیهای ذاتی از والد به فرزند کارکرد دارند.
- در سنتز پروتئین نقش مهمی دارند.
- در جدلهای قانونی از DNA برای حدس و گمان در مورد روابط خانوادگی استفاده میشود.
- همچنین در یافتن مجرم میتواند کاربرد مفیدی داشته باشد.
- در مطالعات مرتبط با ژنتیک و تکامل زیستی نقش پررنگی دارد.
پروتئین
پروتئین بسپاری متشکل از آمینواسیدها است. آمینواسیدها مولکولهایی با گروههای کربوکسیلیک اسیدی و آمینی هستند. بدون وجود پروتئین، لیپیدها و کربوهیدارتها وجود نمیداشتند زیرا آنزیمهایی که برای سنتز آنها استفاده میشود، ساختارهایی پروتئینی هستند.

گفتیم که در این درشت مولکول دو گروه عاملی کربوکسیلیک اسید و آمین وجود دارد. در حالتی که این دو گروه عاملی به وسیله یک گروه کربنی از یکدیگر جدا شده باشند، به پروتئین آلفا میگوییم.
علاوه بر گروه آمینی و کربوکسیلیک اسیدی، این گروه کربنی آلفا نیز به یک هیدروژن و یک گروه دیگر متصل است. این گروه در اندازه و طول میتواند متفاوت باشد. در تصویر زیر این گروه متغیر را با نشان میدهیم. در موجودات زنده ۲۰ آمینواسید در ساختار پروتئینها دخالت دارند و تفاوت آنها تنها در گروه متغیر است. ساختار کلی یک آمینواسید را میتوان بهصورت زیر نشان داد.

در بافت سلولی، درشت مولکول های پروتئینی با سیستم پیچیدهای به نام «ریبوزوم» (Ribosome) به یکدیگر متصل میشوند. بنابراین آمینواسیدها با ترتیبی خاص به یکدیگر متصل میشوند تا پروتئین را به وجود بیاورند درواقع به نحوی متصل هستند که بتوانند اطلاعات ژنتیکی آرانآ را با خود حمل کنند. این ترتیب آمینواسیدها را به نام «ساختار اولیه پروتئین» (Proteine's Primary Sequance) میشناسیم. این ساختار با هیدروژنزدایی از دو آمینواسید مجاور به وجود میآید. بهصورتی که گروه آمینی آمینواسید اول با از دست دادن آب به گروه اسیدی آمینواسید دوم وصل میشود. این پیوند آمیدی جدید را در ساختار پروتئین با نام «پیوند پپتیدی» (Petptide Bond) میشناسیم.
در این ساختار آمینواسیدهای بعدی نیز به ترتیب با اتصال به سر اسیدی ملحق میشوند. توجه داشته باشید که همیشه آمینواسیدهای جدید از سر اسیدی متصل میشوند و این اتصال هیچگاه از سر آمینی صورت نمیگیرد و از آنجا که گروه متغیر میتواند حجیم باشد، آمینواسیدها بهصورت ترانس در ساختار جای میگیرند. ساختار سیس تنها برای یک آمینواسید، «پرولین» (Proline)، مطلوب است. نحوه اضافه شدن آمینواسیدها به ساختار و تشکیل پیوند پپتیدی را میتوانید در تصویر زیر مشاهده کنید.

تفاوت پپتید و پروتئین چیست ؟
پپتید و پروتئین هر دو درشت مولکول هایی هستند که از اتصال آمینواسیدهای مختلف به یکدیگر به وجود میآیند. تنها تفاوت آنها در تعداد آمینواسیدهای موجود در ساختار است. اگر تعداد آمینواسیدهای ساختار کمتر از ۵۰ عدد باشد، به آن پپتید میگوییم. به توالی آمینواسید بالاتر از ۵۰ پروتئین گفته میشود. طولانیترین توالی شناخته شده برای پروتئین حاوی ۳۳۴۲۳ آمینواسید است. در زیر نمونهای از این توالیها را مشاهده میکنید.

ماهیت و عملکرد پپتید و پروتئین توسط ساختار اولیه آن مشخص میشود. ۲۰ آمینواسید در بهوجود آوردن این ساختار نقش دارند.

به دلیل تنوع ساختارهای آمینواسیدی، پروتئینهای بسیار زیادی میتوانند وجود داشته باشند. برای روشن شدن این مسئله مثالی را مورد بررسی قرار میدهیم. اگر بخواهیم پپتیدی متشکل از تنها ۳ آمینواسید داشته باشیم ۸۰۰۰ احتمال متفاوت وجود دارد. در جایگاه هر آمینواسید ۲۰ گزینه متفاوت وجود دارد و میشود.
حالا اگر بخواهیم به همین ترتیب احتمال وجود پپتیدهای ۴۰ آمینواسید را بررسی کنیم، نتیجه خواهد بود. تمام این پپتیدهای احتمالی با توجه به آمینواسیدهایشان ویژگیهای متفاوتی خواهند داشت. بنابراین آشنایی با آمینواسیدهای مختلف و ویژگیهایی که از خود بروز میدهند، خالی از لطف نیست.

همانطور که در تصویرهای بالا مشاهده کردید، میتوان آمینواسیدهای مختلف را با توجه به گروه متغیرشان دستهبندی کرد. آمینواسیدهای ناقطبی و آبگریز در گروه متغیر خود تنها دارای هیدروژن و کربن هستند. آنهایی که دارای گروههای الکترونگاتیوتری مانند اکسیژن و هیدروژن هستند میتوانند پیوند هیدروژنی تشکیل دهند و قطبی هستند. برخی دارای گروه عاملی کربوکسیلیک اسید هستند و خاصیت اسیدی دارند. باقی به دلیل داشتن آمین، میتوانند به عنوان باز عمل کنند. ویژگیهای آمینواسیدها به پروتئین کمک میکند که تا بخورد و ساختار سه بعدی پیدا کند.
این ساختار سهبعدی برای داشتن فعالیت زیستی در پروتئین ضروری است. در بین آمینواسیدهایی که مورد بررسی قرار دادیم، پرولین استثنا است زیرا سر آمینی این آمینواسید در حلقه وجود دارد و همین حلقه باعث میشود که پرولین ترجیح بدهد در یک توالی بهصورت سیس قرار بگیرد. این ویژگی باعث میشود که پروتئینها در محل پرولین خود خم شوند و تغییر مسیر دهند.

آمینواسید نیز دارای ویژگی منحصر بهفردی است. بسیاری از پروتئینهای طبیعی، متیونین را به عنوان اولین آمینواسید خود دارند. آمینواسید سیتئین به دلیل داشتن گروه عاملی تیولی میتواند با سیستئینی دیگر در توالی اکسید شود و پیوند دیسولفیدی ایجاد کند. این پیوند که به پل دیسولفیدی معروف است به تاخوردن پروتئین کمک میکند و پایداری آن را بالا میبرد. این تاخوردگی را میتوانید در تصویر بالا مشاهده کنید.
شکل و عملکرد پروتئین
پیشتر گفتیم که عملکرد یک پروتئین بستگی زیادی به شکل آن دارد. در حالت کلی پروتئینها با چهار ساختار شناخته میشوند.
- ساختار اولیه پروتئین
- ساختار ثانویه پروتئین
- ساختار سوم پروتئین
- ساختار چهارم پروتئین
در بخشهای بعدی به معرفی هر کدام از این ساختارها بهتفضیل خواهیم پرداخت. آگاهی به ویژگیهای هر کدام و تشخیص آنها از یکدیگر به ما در فهم این دسته از درشت مولکول ها کمک بهسزایی میکند.
ساختار اولیه پروتئین
ساختار اولیه هر پروتئین الگوی تاخوردگی آن را مشخص میکند. این ساختار خود خطی است و با تاخوردگی به ساختارهای پیچخورده تبدیل میشود. توالی این ساختار اولیه را میتوانید در تصویر زیر ببینید.

ساختار ثانویه پروتئین
در هر پروتئین قسمتهای کوچک تاخوردگیهای متعددی را متحمل میشوند و این منجر به تولید ساختار ثانویه پروتئین میشود. ساختار ثانویه بستگی به شکل خود دارای دو زیرگروه است.
- «مارپیچ آلفا» (Alpha-Helix)
- «صفحه بتا» (Beta-Pleated-Sheet)
برای حفظ این ساختار سه بعدی برهمکنشهای درونمولکولی بهخصوص پیوند هیدروژنی بین گروههای عاملی آمین و کربونیل ضروری است. هر پیچ ساختار آلفا دارای ۳٫۶ آمینواسید است. در این ساختار گروههای متغیر هر آمینواسید به سمت خارج مارپیچ جهتگیری میکنند.
در ساختار صفحه بتا، بین صفحهها پیوند هیدروژنی از جانب اتمهای زنجیره پلیپپتیدی وجود دارد. در این ساختار گروههای متغیر در بالا و پایین هر صفحه قرار میگیرند و دو صفحه به دو صورت موازی و غیرموازی هستند. همچنین بین نیتروژنهای گروه عاملی آمین با بار جزئی مثبت و اکسیژنهای گروه عاملی کربوکسیلیک اسیدی با بار جزئی منفی، پیوند هیدروژنی وجود دارد. هر دو این ساختارها در بسیاری از پروتئینها مشاهده میشوند و برای حفظ ساختار آنها اهمیت دارند.

پیشتر گفتیم که ساختار ثانویه پروتئینها در حالت صفحه بتا دارای دو نوع موازی و غیرموازی است. در حالت موازی پایانههای اسیدی زیر هم و پایانههای آمینی نیز زیر هم قرار دارند اما در حالت غیرموازی دو صفحه جوری کنار هم قرار گرفتهاند که هر سر آمینی در صفحه زیری خود نزدیک به سر اسیدی باشد و بالعکس. این گفته را میتوانید به خوبی در تصویر زیر مشاهده کنید.

ساختار سوم پروتئین
به شکل سهبعدی کامل پروتئین با در نظر گرفتن تمام زنجیرههای آن، ساختار سوم پروتئین میگویند. این ساختار منحصر بهفرد است و ویژگیهای پروتئین را مشخص میکند. این سختار سهبعدی در وهله اول توسط برهمکنش بین گروههای متغیر آمینواسیدها مختلف سازنده توالی پروتئینی به وجود میآید. ماهیت این گروهها میتواند در تشکیل پیوند هیدروژنی در ساختار ثانویه مشکل ایجاد کند و آن را از بین ببرد. در ادامه مثالی از این موقعیت را مورد بررسی قرار میدهیم. گروههای متغیر در صورتی که باردار باشند و بار آنها همنام باشد، میتوانند یکدیگر را دفع کنند و اگر همنام باشند عکس این اتفاق میافتد، یعنی یکدیگر را جذب میکنند و پیوندی یونی بین آن دو تشکیل میشود.
با ایجاد تاخوردگی در ساختار پروتئین گروههای متغیر آمینواسیدهای غیرقطبی، در قسمت داخلی پروتئین قرار میگیرند، درحالی که گروههای متغیر آبدوست در قسمت خارجی خواهند بود. مورد اول به نام برهمکنش آبگریز مشهور است. همچنین تنها پیوند کووالانسی که طی تشکیل پروتئين به وجود میآید، برهمکنش بین زنجیره جانبی آمینواسید سیستئین با اکسیژن است که منجر به تشکیل پل دیسولفیدی میشود. تمام این برهمکنشهای ضعیف و قوی، ساختار سهبعدی نهایی این درشت مولکول را مشخص میکنند و با از دست رفتن آنها، پروتئین نیز عملکرد خود را از دست خواهد داد.

ساختار چهارم پروتئین
در طبیعت، پروتئینها از چندین «پلیپپتید» (Polypeptide) به وجود میآیند که با نام «زیرواحد» (Subunit) شناخته میشوند. برهمکنش این زیرواحدها باعث به وجود آمدن ساختار چهارم پروتئین میشود. درواقع برهمکنش ضعیف بین این زیرواحدها به تشکیل این ساختار کمک میکند. در این مورد مثالی را بررسی خواهیم کرد.
انسولین که درشت مولکولی کروی است متشکل از پیوندهای هیدروژنی و دیسولفیدی متعددی است که باعث به وجود آمدن آن بهصورت پروتئینی تودهای میشود. این پروتئین در حالت تکپار خود فعال است. انسولین مازاد در بدن بهصورت بسپار که فرمی غیرفعال است، ذخیره میشود تا در زمان نیاز با شکستن و تولید تکپار عملکرد خود را داشته باشد. در تصویر زیر ساختار این پروتئین کروی را مشاهده میکنید.

در بخشهای پیش رو میخواهیم به چند مفهوم مهم در رابطه با این خانواده از درشت مولکول ها بپردازیم.
هیدولیز پروتئین
به فرایندی که طی آن ساختار اولیه پروتئین با افزوده شدن آب، تبدیل به آمینواسیدهای سازنده خود میشوند، هیدرولیز یا آبکافت میگوییم. این مفهوم را میتوانید بهخوبی در تصویر زیر مشاهده کنید. در این فرایند پیوند پپتیدی شکسته میشود و آمینواسیدهای اولیه آزاد خواهند شد.

واسرشتن پروتئین
«واسرشتن» یا «دناتوره شدن» (Denaturation) زمانی اتفاق میافتد که ساختار پیچخورده پروتئین در اثر تغییر دما، تغییر و قرارگیری در معرض مواد شیمیایی، تاخوردگی خود را از دست بدهد و بدون شکسته شدن ساختار اولیه، شکل خود را از دست بدهد. توجه داشته باشید که دناتوره شدن با هیدولز پروتئین متفاوت است زیرا ساختار پروتئین در دناتوره شدن دچار شکستگی نمیشود.
این فرایند معمولا برگشتپذیر است و با از بین رفتن عامل دناتوره شدن، پروتئین ساختار پیشین خود را به دست میآورد و عملکرد عادی خود را خواهد داشت. با اینحال گاهی اوقات این واکنش برگشتناپذیر است و عملکرد پروتئین بهکلی از بین میرود. در بخش بعدی مثالی از دناتوره شدن برگشتناپذیر پروتئین را بررسی خواهیم کرد.

مثالی از دناتوره شدن برگشتناپذیر پروتئین در پختن تخممرغ است. با قرارگیری تخممرغ در آب جوش پروتئین «آلبومین» (Albumin) موجود در سفیده، دناتوره میشود. توجه داشته باشید که نباید دناتوره شدن آلبومین در دمای بالا را به تمامی پروتئینها نسبت بدهیم. مثلا باکتریهای موجود در منابع آب گرم دارای پروتئینهایی هستند که در دمایی نزدیک به نقطه جوش عملکرد خود را حفظ میکنند. همچنین معده انسان که محیطی بسیار اسیدی با پایین است، برای هضم از فرایند دناتوره کردن استفاده میکند. آنزیمهایی که در هضم غذا دخیل هستند در این فضا فعالیت خود را حفظ میکنند و دناتوره نمیشوند.
شپرون چیست ؟
گفتیم که تاخوردگی پروتئین برای عملکرد آن ضروری است و در گذشته این باور وجود داشت که خود پروتئینها در ایجاد این ساختار پیچخورده دخالت دارند. امروزه میدانیم که پروتئینها برای ایجاد تا خوردگی از موادی به نام «شپرون» (Chaperone) کمک میگیرند. شپرونها از تجمع پلیپپتیدها جلوگیری میکنند و بعد از ایجاد تاخوردگی در پروتئین از آن جدا میشوند.
نکات تکمیلی پروتئین
در این بخش به برخی از مهمترین کاراییهای این درشت مولکول های زیستی میپردازیم.
- به عنوان آنزیم در افزایش سرعت واکنشهای زیستی کاربرد دارند.
- بخشی جداییناپذیر در فرایندهایی هستند که منجر به شکست سلول میشود.
- در سیگنالدهی سلولی و انتقال مولکول از نقطهای به نقطه دیگر کاربرد دارند.
- در شرایط حاد به عنوان منبعی برای تغذیه سلولها خواهند بود.
لیپید
لیپیدها با نام متداول چربی، درشت مولکول هایی «آبگریز» (Hydrophobic) هستند، به این معنی که در آب حل نمیشوند. این گروه از درشت مولکول ها شامل «تریگلیسیرید» (Triglyceride)، «کارتنوئید» (Cartenoid)، «فسفولیپید» (Phospholipid) و «استروئید» (Steroid) هستند.
از کارکرد لیپیدها میتوان به کمک به تشکیل دیواره سلولی و تشکیل هورمونها اشاره کرد. همچنین لیپیدها در بدن به عنوان سوخت ذخیرهای باقی میمانند.

لیپیدها بهطور عمده از دو عنصر کربن و هیدروژن تشکیل میشوند و همین دلیل آبگریز بودن آنها است. همچنین این درشت مولکولها میتوانند دارای مقادیر جزئی از اکسیژن، نیتروژن، گوگرد و فسفر باشند. لیپیدها در عملکرد ارگانها نقش بهسزایی دارند، به برخی از آنها اشاره خواهیم کرد.
- لیپیدها منبع غذایی غنی هستند.
- مولکولهایی هستند که انرژی را در خود ذخیره میکنند.
- در ساختار غشا حضور دارند.
- در سیگنالدهی شیمیایی نقش دارند.
لیپیدها ساختارهای مهمی را تشکیل میدهند. در ادامه میخواهیم برخی از مهمترین این ساختارهای لیپیدی را مورد بررسی قرار دهیم.
اسید چرب و تریگلیسیرید
«اسیدهای چرب» (Fatty Acid) لیپیدهایی حاوی زنجیرههای بلند کربوهیدراتی با پایانه گروه عاملی کربوکسیلیک اسید هستند. به خاطر همین زنجیره بلند کربوهیدراتی، اسیدهای چرب غیرقطبی و آبگریز هستند.
به اسیدهای چربی که فقط حاوی پیوندهای یگانه باشند، «اسید چرب اشباع» (Saturated Fatty Acid) میگویند زیرا بیشترین تعداد اتمهای هیدروژن ممکن را دارند و اشباع شدهاند. در جدول زیر تعدادی از اسیدهای چرب اشاع را مشاهده میکنید.
| نام اسید چرب اشباع | ساختار شیمیایی | نقطه ذوب (سانتیگراد) |
| لوریک اسید | ۴۴ | |
| پالمیتیک اسید | ۶۳ | |
| استئاریک اسید | ۷۰ |
اسیدهای چربی که حداقل یک پیوند دوگانه در ساختار خود داشته باشند، اشباع نیستند زیرا دارای هیدروژنهای کمتری هستند و به خانواده «اسیدهای چرب غیراشباع» (Unsaturated fatty Acid) تعلق دارند. در جدول زیر تعدادی از اسیدهای چرب غیراشباع را بههمراه نقطه ذوب آنها مشاهده میکنید.
| نام اسید چرب غیراشباع | ساختار شیمیایی | نقطه ذوب (سانتیگراد) |
| اولئیک اسید | ۱۶ | |
| لینولئیک اسید | ۵- | |
| لینولنیک اسید | ۱۱- | |
| آراشیدونیک اسید | ۵۰- |
اسیدهای چرب اشباع ساختاری خطی و اسکلت کربنی منعطفی دارند در حالی که اسیدهای چرب غیراشباع دارای پیچ و تابهایی هستند. دلیل این تفاوت در پیوندهای کربن-کربن دوگانه است که ساختاری صلب دارد و امکان پیچ خوردن را از بین میبرد. این تفاوتها باعث تفاوت در ویژگی لیپیدهایی میشوند که دارای اسید چرب اشباع و غیراشباع هستند. بهطور مثال لیپیدهای حاوی اسیدهای چرب اشباع در دمای اتاق جامد هستند اما لیپیدهای حاوی اسیدهای چرب غیراشباع در شرایط مشابه مایع هستند.

مقایسه نقطه ذوب اسید چرب اشباع با اسید چرب غیراشباع
با توجه به دو جدول بالا متوجه میشوید که نقطه ذوب اسیدهای چرب غیراشباع از اسیدهای چرب اشباع بیشتر است. برای پی بردن به چرایی این قضیه باید به ساختار هندسی این دو گروه نگاه کنیم. زاویه پیوندی چهاروجهی روی کربن باعث به وجود آوردن حالتی در اسیدهای چرب اشباع میشود که بهصورت خطی و زیگزاگی باشند. بنابراین این مولکولها متراکم و فشرده به هم هستند و این باعث میشود که نیروی بینمولکولی و در نتیجه نقطه ذوب بالاتر باشد.

از طرف دیگر، وجود یک یا چند پیوند دوگانه در اسید چرب غیراشباع باعث میشود که در هر محل پیون دوگانه، مولکول کمی خم شود. این پیوندهای دوگانه موجود در اسیدهای چرب غیراشباع در بیشتر موارد بهصورت ایزومر «سیس» (Cis) وجود دارند و توده متراکمی را تشکیل نمیدهند. بنابراین به دلیل کاهش نیروی بینمولکولی، نقطه ذوب در اسیدهای چرب غیراشباع نسبت به اسیدهای چرب اشباع پایینتر است.
در لیست زیر مقدار و نوع اسیدهای چرب اشباع و غیراشباع را در برخی منابع غذایی مشاهده میکند.
منشا حیوانی
- کره حیوانی: پالمتیک اسید (۲۹٪)، استئاریک اسید (۹٪)، اولئیک اسید (۲۷٪)، لینولئیک اسید (۴٪)، اسیدهای چرب دیگر (۳۱٪)
- گوشت گاو: پالمتیک اسید (۳۲٪)، استئاریک اسید (۲۵٪)، اولئیک اسید (۳۸٪)، لینولئیک اسید (۳٪)، اسیدهای چرب دیگر (۲٪)
منشا گیاهی
- روغن ذرت: پالمتیک اسید (۱۰٪)، استئاریک اسید (۴٪)، اولئیک اسید (۳۴٪)، لینولئیک اسید (۴۸٪)، اسیدهای چرب دیگر (۴٪)
- سویا: پالمتیک اسید (۷٪)، استئاریک اسید (۳٪)، اولئیک اسید (۲۵٪)، لینولئیک اسید (۵۶٪)، اسیدهای چرب دیگر (۹٪)
- بادام زمینی: پالمتیک اسید (۷٪)، استئاریک اسید (۵٪)، اولئیک اسید (۶۰٪)، لینولئیک اسید (۲۱٪)، اسیدهای چرب دیگر (۷٪)
- زیتون: پالمتیک اسید (۶٪)، استئاریک اسید (۴٪)، اولئیک اسید (۸۳٪)، لینولئیک اسید (۷٪)
برای درک بهتر دو ماده غذایی روغن زیتون و کره را با یکدیگر مورد مقایسه قرار میدهیم.

گفتیم که اسیدهای چرب اشباع در دمای اتاق بهصورت جامد حضور دارند اما اسیدهای چرب غیراشباع به دلیل داشتن پیوند دوگانه در دمای مشابه مایع هستند. به توجه به اطلاعات بالا متوجه میشویم که کره بیشتر از اسیدهای چرب اشباع تشکیل شده است بنابراین در دمای اتاق جامد است اما روغن زیتون اینطور نیست. این ماده به دلیل داشتن مقدار زیادی اسیدچرب غیراشباع در دمای اتاق به فرم مایع حضور خواهد داشت.
«تریگلیسیرید» (Triglyceride) یا «تری آسیل گلیسرول» (Triacylglucerol) زمانی به وجود میآید که سه اسید چرب به یک مولکول گلیسرول متصل شوند. تریگلیسیریدها سازنده «آدیپوز» (Adipose)، چربی بدن و «سبوم» (Sebum) چربی پوست، هستند.
تریگلیسیریدها در متابولیسم بدن تاثیر بهسزایی دارند زیرا نسبت به کربوهیدارتها و پروتئینها، منبع بسیار بهتری برای انرژی هستند. در شکل زیر نحوه تشکیل این چربی طبیعی نشان داده شده است. طی این واکنش آبزدایی، سه مولکول آب از دست میرود و مولکول گلیسرول و سه زنجیره اسید چرب به هم متصل میشوند.

اسید چرب مضر و مفید
در اواخر ۱۹۶۰ فرضیهای مطرح شد مبنی بر اینکه مصرف اسید چرب اشباع منجر به بالا رفتن سطح «لیپوپروتئين کمچگالی» (Low Density Lipoprotein) میشود که به نظر کلسترول مضری بود. این قضیه از طریق نازک کردن عروق باعث سکته قلبی میشد و در نهایت نیز چربیها به دو دسته مضر و مفید تقسیمبندی شدند. در این دستهبندی اسیدهای چرب اشباع مضر در نظر گرفته میشدند و اسیدهای چرب غیراشباع، مفید. در سالهای بعد دانشمندان تحقیقات زیادی را روی این فرضیه انجام دادند و متوجه شدند که هیچ ارتباط مستقیم بین مصرف اسیدهای چرب اشباع و سکته قلبی وجود ندارد، بنابراین این فرضیه منسوخ در نظر گرفته شد. مصرف اسیدهای چرب فواید بسیار مهمی بر بدن انسان دارد که در لیست زیر برخی از آنها را آوردهایم.
- سلامت سلولها به آنها بستگی دارد.
- به تقویت ذهن کمک میکنند.
- مصرف ویتامینهای محلول را در بدن تسهیل میکنند.
- از ایجاد «ترومای بلانت» (Blunt Trauma) در اندامهای بالشی بدن جلوگیری میکند.
هیدروژناسیون اسیدهای چرب غیراشباع و اسیدهای چرب ترانس
واکنشهای هیدروژناسیون ساده توانایی این را دارند که اسیدهای چرب غیراشباع را به اسیدهای چرب اشباع تبدیل کنند. توجه داشته باشید که هیدروژناسیون آلکن، محصول آلکانی به دست میدهد. صورت کلی این واکنش را میتوانید در زیر مشاهده کنید.
معمولا از روغنهای گیاهی با عنوان «چند غیراشباعی» (Polyunsaturated) نام برده میشود و منظور این است که تعداد پیوندهای دوگانه آنها بیش از یکی است. این روغنها میتوانند در اثر هیدروژناسیون از مایع تبدیل به جامد شوند. در تصویر زیر واکنشی که طی آن اسید چرب غیراشباع به اسید چربی اشباع تبدیل میشود را مشاهده میکنید. در این واکنش پیوند دوگانه تبدیل به پیوند یگانه شده است.

روغنهای گیاهی که بهصورت جزئی هیدروژنه شده باشند، یعنی تنها تعدادی از پیوندهای دوگانهشان را از دست داده باشند، با افزایش نقطه ذوب روبرو میشوند. درواقع درجه هیدروژناسیون، حالت نهایی آن را در محیط مشخص میکند.
اسید چرب ترانس
یکی از بزرگترین دغدغهها در واکنش هیدروژناسیون اسیدهای چرب غیراشباع، تولید اسید چرب ترانس است. اسیدهای چرب نتیجه واکنشی جانبی در هیدروژناسیون کاتالیزوری هستند. همانطور که پیشتر اشاره کردیم، اسیدهای چرب غیراشباع بهصورت طبیعی به فرم ایزومری ترانس حضور دارند. طبق تعریف ایزومرها، مولکولهایی با فرمول مولکولی برابر اما نحوه اتصال متفاوت هستند. در ایزومر سیس هیدروژنها در یک سمت قرار دارند و در فرم ایزومری ترانس در دو سمت مختلف. به دلیل انرژی که در واکنش هیدروژناسیون وارد محیط میشود، توانایی این را دارد که پیوندهای دوگانه اسیدهای چرب غیراشباع را از فرم سیس به ترانس تبدیل کند، یعنی یکی از هیدروژنها از سمتی به سمت دیگر منتقل میشود. بدن انسان توانایی تشخیص این اسید چرب غیراشباع ترانس را ندارد.

با وجود اینکه تفاوت این دو تنها در سیس و ترانس بودن پیوندهای دوگانه است، بسیار متفاوت از هم هستند. بزرگترین نقطه ضعف اسید چرب ترانس این است که باعث افزایش کلسترول مضر میشود. برخی از مواد خوارکی که در آنها اسید چرب ترانس وجود دارد، شامل موارد زیر هستند.
- دونات
- شیرینی دانمارکی
- مرغ سوخاری
- سیبزمینی سرخکرده
فسفولیپید و غشای بیولوژيکی
تریگلیسیریدها جزو لیپیدهای ساده هستند زیرا تنها از دو عضو تشکیل میشوند: اسید چرب و گلیسرول. لیپیدهای پیچیده از حداقل ۳ عضو تشکیل میشوند، بهطور مثال گروه فسفاتی (فسفولیپید) یا زنجیره کربوهیدراتی (گلیکولیپید). زمانی که دو اسید چرب، به یک مولکول گلیسرول متصل شوند و ظرفیت سوم این مولکول توسط گروهی فسفاتی اشغال شود، یک فسفولیپید به وجود میآید.
این دو اسید چرب میتوانند هر دو اشباع باشند یا هر دو غیراشباع باشند. در حالت سوم ممکن است یکی از اسیدچربها اشباع و دیگری غیراشباع باشد.

تریگلیسیریدها به دلیل ناقطبی بودن اسیدهای چرب موجود در ساختار خود، همه ناقطبی هستند و تمایلی به مولکولهای آب ندارند، در نتیجه آبگریز هستند. از طرفی فسفولیپیدها آبدوست هستند زیرا دارای گروه فسفاتی با بار منفی هستند. این گروه فسفاتی توانایی این را دارد که با آب جاذبهای قوی ایجاد کند. به قسمت آبدوست فسفولیپید بهاصطلاح سر قطبی گفته میشود. به مولکولهایی مانند فسفولیپیدها که دارای یک بخش آبگریز و یک بخش آبدوست هستند، «آمفی پاتیک» (Aphipathic) گفته میشود.
این ویژگی به فسفولیپیدها عملکرد منحصر بهفردی را در محیطهای آبی میبخشد. همانطور که پیشتر گفتیم این ترکیبات دارای ساختاری با یک سر آبدوست و یک سر آبگریز هستند. از آنجا که فسفولیپیدها طول زیادی دارند، سرهای آبگریز با قدرت زیادی به یکدیگر متصل هستند. با توجه به مواردی که گفتیم فسفولیپیدها ساختاری کروی به خود میگیرند. با نگاه به این درشت مولکول متوجه میشویم که دمهای غیرقطبی در تماس با سطح نیستند. در این حالت فسفولیپیدها به سه حالت میتوانند وجود داشته باشند.
- لیپید کروی یکلایه
- لیپید کروی دولایه
- لیپید صفحهای دولایه
غشای سلولی بسیاری از ارگانیسمها و اجزای درونسلولی از لیپید صفحهای دولایه تشکیل میشود.

در بخشهای پیش رو میخواهیم دو فسفولیپید مهم و پرکاربرد را مورد بررسی قرار دهیم.
لسیتین
این مولکول یکی از رایجترین اعضای خانواده فسفولیپیدها است و در زرده تخممرغ، جوانه گندم و دانه سویا یافت میشود. این ماده را از پس از استخراج از دانه سویا به عنوان «عامل امولسیون کننده» (Emulsifying Agent) به مواد غذایی اضافه میکنند. از آنجا که لسیتین دارای هر دو ویژگی قطبیت و غیرقطبیت است میتواند با مخلوط کردن چربی و روغن با مواد آبی، در این زمینه به کار رود.
همچنین این ماده در لیپید دولایهای غشای سلول نقش پررنگی را ایفا میکند. در ساختار لسیتین نمک آمونیوم کولین به کمک پیوندی استری به گروه فسفات متصل است و نیتروژن آن دارای بار مثبت و ۴ گروه متیلی است.

سفالین
سفالین، فسفوگلیسریدی حاوی اتانول آمین یا آمینواسید سرین متصل به گروه فسفاتی است. اسیدهای چرب متنوعی در باقی ساختار آن مشاهده میشود. این درشت مولکول در غشای سلولی بهخصوص در مغز یافت میشود. همچنین وظیفه ایجاد لخته در خون نیز با این ماده است.

ایزوپرونوئيد و استرول
«ایزوپرنوئیدها» (Isopernoid) لیپیدهایی شاخهای هستند که با نام «ترپنوئید» (Terpenoid) نیز از آنها یاد میشود. این ترکیبات با تغییراتی روی مولکول «ایزوپرن» (Isoprene) به وجود میآید.این لیپیدها نقش بسیار بزرگی در فیزیولوژی حیوانات و گیاهان دارند. همچنین در بسیاری از ترکیبهای دارویی مانند «کاسپایسین» (Caspaicin)، رنگدانهها مانند «زانتوفیل» (Xanthophyl) بتا کاروتن (Beta-Carotene) و رایحههایی مانند «لیمونین» (Limonene)، «منتول» (Menthol) و کافور حضور دارند.

همچنین میتوان در روغنهای طبیعی و «مومها» (Wax)، ایزوپرنوئیدهای بلند زنجیر را مشاهده کرد. مومها معمولا در برابر نفوذ آب مقاوم و در دمای اتاق سفت هستند اما با بالا رفتن دما، موم نرم میشود و در دماهای خیلی بالا میتواند بهصورت مایع حضور داشته باشد. در بدن انسان اصلیترین موم توسط «غدد چربی فولیکولهای مو» (Sebaceous glands of hair follicles) در پوست تولید میشود و منجر به ترشح مادهای به نام «سبوم» (Sebum) میشود. بخش عمده این ماده را تریگلیسیرید، مومهای استری و هیدروکربن «اسکوالین» (Squalene) است. باکتریهای بسیاری روی سطح پوست قرار دارند که از لیپیدها تغذیه میکنند. یکی از مهمترین این باکتریها «پروپیونی باکتریوم» (Propionibacterium) آکنه است. این باکتری از این لیپیدها برای تولید اسیدهای چرب کوتاه زنجیر استفاده میکند و به این طریق در ایجاد آکنه روی پوست دخالت دارد.
لیپیدهای پیچیده دیگری نیز وجود دارند که حلقوی هستند و در غشای سلولی یافت میشوند و برخیشان عملکرد هورمونی دارند. این لیپیدها را با عنوان «استروئید» (Steroid)میشناسیم و معروفترین استروئید استرولها هستند که درواقع استروئیدهایی حاوی گروه عاملی میباشند.
این درشت مولکول ها بهطور عمده آبگریز هستند اما دارای گروه آبدوست هیدروکسیل هستند. متداولترین استرولی که در بافت بدن حیوانات دیده میشود، کلسترول نام دارد. ساختار کلسترول دارای ۴ حلقه است که یکی از این حلقهها در خود پیوندی دوگانه دارد. کلسترول باعث استحکام غشای سلولی در «یوکاریوتها» (Eukaryote) و باکتریها میشود.

پروستاگلاندین
پروستاگلاندینها مانند هورمون هستند، زیرا به عنوان پیامرسان شیمیایی عمل میکنند اما برخلاف هورمونها، این درشت مولکول این کار را در همان محلی که سنتز میشوند، انجام میدهند و به نقاط دیگر منتقل نمیشوند.
این درشت مولکول، کربوکسیلیک اسیدی غیراشباع است که از اسکلتی با ۲۰ کربن تشکیل شده است و از نظر بیوشیمیایی از اسید چرب آراشیدونیک اسید به دست میآید. همچنین این درشت مولکول در ساختار خود یک حلقه ۵ عضوی دارد. ساختار ویژه آراشیدونیک اسید توالی پیوندهای دوگانه را جوری کنار هم قرار داده است که به تشکیل این حلقه کمک میکند. ساختار این درشت مولکول و حلقه ۵ عضوی آن را میتوانید در تصویر زیر مشاهده کنید.

پروستاگلاندین میتواند روی حلقه خود دارای پیوندی دوگانه، گروه عاملی کتون یا الکل باشد که یکی از آنها را در تصویر بالا آوردیم.
عملکرد پروستاگلاندین
در این بخش میخواهیم به برخی از مهمترین تاثیرات فیزیولوژی این ماده در بدن انسان بپردازیم.
- این درشت مولکول فعالساز پاسخ التهابی، درد و تب است. زمانی که بافتی در بدن آسیب ببینید، گلبولهای سفید خون برای به حداقل رساندن آسیب به محل میروند. در نتیجه این موضوع، پروستاگلاندین تولید میشود.
- وقتی رگ خونی دچار آسیب شود، لخته خون ایجاد میشود. نوعی از پروستاگلاندینها با نام ترومبوکسان تولید لخته از پلاکتهای خود را تسریع میکند.
- عکس عملکرد بالا نیز ممکن است. یعنی کمک به تخریب لختههای خونی که باعث ایجاد دردسر میشوند.
- برخی پروستاگلاندینها در القای زایمان به افراد باردار کمک میکند. این فرایند با ایجاد انقباض در رحم انجام میشود.
- پروستاگلاندین در اندامهای دیگر، مانند اندامهای گوارشی، نیز حضور دارند و باعث افزایش جریان خون در کلیه میشود.
در بخش قبل از ایجاد التهاب، درد و تب توسط پروستاگلاندین صحبت کردیم و این ما را یاد عملکرد «آسپرین» (Aspirin) میاندازد. آسپرین با مسدود کردن آنزیمی به نام «سیلکلواکسیژناز» (COX-1 و COX-2) که باعث حلقهزایی و افزودن اتم اکسیژن به آراشیدونیک اسید و تبدیل آن به پروستاگلاندین میشود، از فعالیت آن جلوگیری میکند. گروه استیل آسپرین هیدرولیز میشود و به عنوان استر به گروه الکلی سرین موجود در ساختار آنزیم، متصل میشود. این باعث مسدود شدن کانال آنزیم میشود و آراشیدونیک اسید نمیتواند وارد سایت فعال آن شود و با این کار سنتز پروستاگلاندین مختل نمیشود. این اتفاق تا حدی از درد و تب نیز میکاهد. همچنین تصور میرود آسپرین از سنتز پروستاگلاندین و ایجاد لخته نامطلوب در رگهای خونی نیز جلوگیری میکند اما مصرف آسپرین میتواند در زمان وارد شدن جراحت به بدن، میزان خونریزی را بسیار بیشتر و شدیدتر کند.

واکنش درشت مولکول ها
درشت مولکول ها با یکدیگر و با مولکولهای کوچک دیگر وارد واکنش میشوند و در این فرایند دستخوش تغییراتی خواهند شد. در ادامه به برخی از این برهمکنشها میپردازیم.
برهمکنش شامل پروتئین
پروتئینها درشت مولکولی با ۴ سطح ساختاری هستند. در این بخش میخواهیم به برهمکنشهایی بپردازیم که این خانواده از درشت مولکول ها را به خود درگیر میکند.
اتصال غیرپروتئین به پروتئين
به درشت مولکول حاصل از اتصال غیر مولکول غیرپروتئین به پروتئین، «پروتئین مزدوج» (Conjugated Protein) گفته میشود. این بخش جدید اضافه شده به پروتئین میتواند نقش زیستی از خود ایفا کند و در عملکرد پروتئین تاثیر مستقیمی داشته باشد. همچنین میتواند با تغییر شکل پروتئین نیز اثرگذار باشد. در زیر تعدادی از ترکیبهایی که میتوانند به پروتئین متصل شوند و تغییراتی را ایجاد کنند، میبینید.
- «هم» (Heme)
- لیپید
- کربوهیدرات
- یون فلزی
- فسفات
بخشهای مختلف پروتئين حاصل را با نامهایی از یکدیگر مجزا میکنند. به جدول زیر دقت کنید.
| نام | تعریف |
| آپوپروتئین (Apoprotein) | پروتئین ابتدایی مجزا |
| گروه پروستتیک (Prosthetic Group) | بخش غیرپروتئينی مجزا |
| پروتئين مزدوج | مجموع پروتئین ابتدایی + بخش غیرپروتئينی |
پروتئین فلزی
«پروتئين فلزی» (Metalloprotein) گروهی از پروتئینهای هستند که حاوی فلزهای زیر میباشند.

این فلزها میتوانند پیچیدههای کوئوردیناسیونی تشکیل بدهند زیرا میتوانند از اتمهایی با زوج یون غیراشتراکی، الکترون بپذیرند. این الکترون اوربیتال ظرفیت فلز مانند را پر میکند. همچنین برخی از این فلزها به راحتی وارد واکنش اکسایش-کاهش میشوند. به مثال زیر توجه کنید.
در ادامه به برخی ویژگیهای پروتئین فلزی ایجاد شده، اشاره میکنیم.
- در این ساختار پروتئین به عنوان «لیگاندی چنددندانه» (Polydentate Ligand) ایفای نقش میکند و با ایجاد پیچیده به کمک فلز به پایداری ساختار سوم پروتئین کمک میکند.
- فلز میتواند به اتصال پروتئین به مولکولها دیگر کمک کند.
- فلز موجود درعملکرد پروتئین موثر است، مثلا میتواند باعث فعالسازی سطح سوبسترا شود. زمانی که فلز از سوبسترا زوج یون میپذیرد، سوبسترا به دلیل کمبود الکترون فعالتر میشود.
- گفتیم که فلزها میتوانند بهراحتی وارد واکنش اکسایش-کاهش شوند. گاهی اوقات میتوانند این واکنش را بهصورت مستقیم و متصل به پروتئین با عوامل زیستی انجام دهند.
لیپوپروتئين
پروتئینها میتوانند با برهمکنش آبگریز به لیپیدها متصل شوند. این ترکیبها ساختاری شبیه به «میسل» (Micelle) دارند، یعنی دارای سر قطبی و دم آبگریز هستند. در آب سر قطبی به سمت بیرون است و دم غیرقطبی در مرکز درشت مولکول قرار میگیرد.
این ترکیب تفاوت عمدهای با پروتئین مجزا دارد، به دلیل وجود سطح باردار، تمایل دارد به سطح دیگری متصل شود. غلظت لیپوپروتئین در زمان وجود عفونت برای افزایش محافظت، افزایش پیدا میکند. این ترکیب به اندوتوکسین باکتری میچسبد و با اتصال میتواند عملکرد بسیاری از ویروسها را خنثی کند.
پروتئینهای غشایی ساختاری شبیه به لیپوپروتئینها دارند زیرا در مناطق استراتژیک خود دارای آمینواسید غیرقطبی هستند و این به پروتئین قابلیت برهمکنش با لیپید غشایی را میدهد.
گلیکوپروتئين
«گلیکوپروتئين» (Glycoprotein) پروتئينی است که گروه پروستتیک آن کربوهیدرات است. این کربوهیدرات همراه میتواند حاوی ۱ تا ۳۰ واحد مونوساکاریدی چه بهصورت شاخهای چه خطی باشد و با پیوندی کووالانسی به آپوپروتئین وصل است. این دو مولکول به سه روش مختلف با هم همراه میشوند که در ادامه به بررسی آنها خواهیم پرداخت.
- اتصال از اتم N (نوع اول): در این حالت ان-استیل گلوکزآمین (قندی با یک گروه آمین استیله به جای گروه هیدروکسیل) در قسمت کاهشی زنجیره کربوهیدارتی به نیتروژن آمید آمینواسید آسپارژین متصل میشود. این اتصال زمانی صورت میگیرد که توالی آمینواسیدی بهصورت یا وجود داشته باشد. در این توالی میتواند هر آمینواسیدی باشد. توجه داشته باشید که آسپارژین به هر حالت دیگری حضور داشته باشد، فاقد توانایی اتصال از این نوع است.
- اتصال از اتم O (نوع دوم): در این نوع اتصال قسمت کاهشی زنجیره جانبی به (معمولا بهصورت ان-استیل آمین) به هیدروکسیل آمینواسید سرین یا ترئونین متصل است.
- اتصال از اتم O (نوع سوم): در این نوع اتصال قسمت کاهشی زنجیره جانبی که در اینجا نیز معمولا ان-استیل آمینه است به هیدوکسیل هیدروکسی لایزین در کلاژن متصل میشود. ( هیدروکسی لایزین از لایزین در ساختار کلاژن بعد از سنتز آن به وجود میآید.)
درشت مولکولها در موارد دیگری نیز میتوانند با درشت مولکولهای دیگر همراه شوند. برای مثال پروتئین و نوکلئيک اسید این توانایی را دارند.
مثال هایی از درشت مولکول ها
در این بخش میخواهیم برخی از درشت مولکول هایی که پرکاربرد و شناختهشده هستند را مورد بررسی قرار دهیم.
الیاف مصنوعی
«نایلون» (Nylon)، «ریون» (Rayon) و «اسپندکس» (Spandex) همگی درشت مولکول هستند و طی مراحل خاصی تولید میشوند.
- ابتدا مونومرهای سازنده درشت مولکول ها تولید میشوند.
- در مرحله دوم بعد از تشکیل بسپار، از غشایی عبور داده میشوند تا قطر مناسب مورد نیاز به دست آید. به این مرحله بهاصطلاح نخریسی میگویند.
انتقال ژنتیکی
DNAها متشکل از نوکلئیک اسیدهایی هستند که ویژگیهای ژنتیکی را در خود حمل میکنند. در فرایند تقسیم سلولی DNAها کلیت خود را حفظ نمیکنند و نوکلئوتیدهای بهجا مانده از آنها وظیفه انتقال اطلاعات را به عهده دارد.
کاربرد صنعتی درشت مولکول ها
درشت مولکول های زیادی وجود دارند که در صنعت پرکاربرد و حیاتی هستند. از این مورد میتوان به پلاستیک، الیاف و «الاستومر» (Elastomer) میشوند. الاستومر درشت مولکولی است که منعطف است و حالت کشسانی دارد. به همین دلیل از الاستومرها در تولید کش استفاده میشود. درواقع ساختار آنها بهگونهای است که به راحتی کشیده میشود و قابلیت این را دارد که بعد از رها شدن به حالت اولیه خود برگردد.
الیاف مصنوعی مواد مورد نیاز در صنعت پوشاک هستند و در تمامی آیتمها مانند تیشرت و کفش به کار گرفته میشود. در کنار الیاف مصنوعی، الیافی طبیعی نیز وجود دارند که شامل پشم، نخ و ابریشم هستند. بنابراین تعداد زیادی از لوازم مورد استفاده در روزمره ما از درشت مولکول ها ساخته میشوند.
کروماتوگرافی اندازه طردی
«کروماتوگرافی اندازه طردی» (Size Exclusion Chromatography) نوعی از روش جداسازی کروماتوگرافی است که ذرات را بر اساس تفاوت در اندازه و هندسه آنها جدا میکند. در این روش گاهی میتوان از تفاوت جرم مولکولی ترکیبهای مختلف نیز بهره برد.
این روش بهطور عمده برای جداسازی درشت مولکول هایی مانند پروتئین، آنزیم، «پادتن» (Antibody)، بسپارهای صنعتی و نوکلئیک اسیدهای RNA و DNA مورد استفاده قرار میگیرد. در تصویر زیر نمایی از فرایند انجام این نوع کروماتوگرافی را مشاهده میکنید.

اساس این روش جداسازی مولکولها با اندازههای مختلف با گذراندن آن ها از بستری متخلخل است. این بستر میتواند بسته به نمونه موردنظر تغییر کند تا مناسب جداسازی باشد. در این روش ابتدا مولکولهای کوچک وارد حفرههای بستر میشوند و بهاصطلاح «زمان بازداری» (Retention Time) بیشتری دارند. در ادامه جداسازی طی دو مرحله زیر صورت میگیرد.
- مولکولهای بزرگ که توانایی عبور از حفره را ندارند، سریع شسته شده و از ستون خارج میشوند.
- سپس مولکولهای کوچک که در حفرهها نفوذ کردهاند، بهآرامی شسته میشوند و جداسازی صورت میگیرد.
همانطور که مشاهده کردید مولکولهای بزرگ نمونه در فرایند شویش زودتر از بستر خارج میشوند. بنابراین زمان بازداری آنها از مولکولهای کوچک، کمتر است. بنابراین بین اندازه مولکول و زمان بازداری رابطه عکس وجود دارد.
کاربرد کروماتوگرافی اندازه طردی
این کروماتوگرافی روشی بسیار مفید و کارامد برای جداسازی درشت مولکول است. در زیر لیستی از مهمترین ترکیبهایی که به این روش جداسازی میشوند را آوردهایم.
- قند
- پلیپپتید
- ویروس
- پروتئین
- آنزیم
- هورمون
- پادتن
- نوکلئیک اسید
با اینحال کاربرد عمده این روش در جداسازی پروتئینها و بسپارهای محلول در آب است. همچنین از این رش برای بررسی وجود و ویژگیهای ترکیبهای آلی موجود در آب استفاده میشود.
مثال و حل تمرین
در این بخش میخواهیم برای درک بهتر موضوع و سنجش میزان آموزش به بررسی چند مثال و تمرین بپردازیم.
تمرین اول
در شیمی کربوهیدارتها معمولا حلقههای فورانوزی و پیرانوزی را بهصورت تخت و به کمک طرح هاوارث رسم میکنند. به دو تصویر زیر توجه کنید.

اگر استخلافهای روی هر کربن را رسم کنیم، نتیجه بهصورت زیر خواهد بود.

مرحله اول: در این مثال میخواهیم بدانیم چطور میتوانیم با در دست داشتن تصویر فیشر یک قند، فرم حلقوی آن را تولید کنیم و با طرح هاوارث رسم کنیم. مولکول موردنظر بهصورت زیر است.

مرحله دوم: ابتدا نیاز داریم که کربنهای ساختار را به ترتیب عددگذاری کنیم.

مرحله سوم: برای داشتن حلقه پیرانوزی اکسیژن روی کربن شماره ۵ باید با پیوندی یگانه به کربن شماره ۱ متصل شود. این را با رسم فلشی آبی در تصویر زیر نشان دادهایم.

مرحله چهارم: در این مولکول، کربن ۱ پشت صفحه کاغذ و گروه هیدروکسیل روی کربن ۵ در جلوی صفحه قرار دارد. برای اینکه حلقه پیرانوزی مسطح باشد، این دو باید در یک جهت باشند، یعنی یا هر دو روی صفحه باشند یا هر دو پشت آن قرار بگیرند. کربن شماره ۵ مرکز کایرال است. برای اینکه جایگاه گروه هیدروکسیل روی کربن را با گروه متصل به کربن ۵ یعنی عوض کنیم بهطوری که در این فرایند پیکربندی مطلق کربن شماره ۵ تغییر نکند ، نیاز است که سه لیگاند متصل به آن یعنی ، و را بدون دست زدن به لیگاند چهارم در جهت عقربههای ساعت بچرخانیم. در تصویر زیر این را نشان دادهایم.

مرحله پنجم: این دو ساختار هر دو نشاندهنده دی-گلوکز هستند اما در ساختار سمت راست برخلاف ساختار دیگر، گروه هیدروکسیل کربن شماره ۱ و کربن شماره ۵ در یک سمت قرار دارند. در مرحله بعد قوانین رسم فرم فیشر را نادیده میگیریم و مولکول را ۹۰ درجه در جهت موافق حرکت عقربههای ساعت میچرخانیم. در تصویر زیر این موضوع نشان داده شده است.

مرحله ششم: در مرحله بعد مولکول را در محور عمودی بهصورتی که در تصویر زیر نشان داده شده است، رسم میکنیم.

مرحله هفتم: استخلافهای موجود روی هر کربن را روی مولکول مرحله قبلی اضافه کنید. توجه داشته باشید که موقعیت لیگاندهایی که در مرحله پنجم رو به بالا بودند در اینجا نیز به به سمت بالا رسم میشوند. همین موضوع در مورد استخلافهای رو به پایین در مرحله پنجم نیز صدق میکند.

مرحله هشتم: در این مرحله برای حلقهزایی باید اتم هیدروژن و اکسیژن روی کربن شماره ۱ و هیدروژن روی اکسیژن متصل به کربن ۵ را حذف کنیم و این دو اتم را به یکدیگر متصل کنیم.

مرحله نهم: با توجه به کربن ۱ متوجه میشویم که دارای دو ظرفیت خالی است. ابتدا دو پیوند یگانه برای آن رسم میکنیم.

مرحله دهم: در موقعیت بالا یک اتم هیدروژن و در موقعیت پایین یک گروه هیدروکسیلی وصل میکنیم. نتیجه بهصورت زیر خواهد بود.

مرحله یازدهم: جای دو روی کربن ۱ را با یکدیگر عوض میکنیم.

تصویر مرحله دهم و یازدهم طرح هاوارث حلقه پیرانوزی دی-گلوکز است. اگر کربنی که گروه هیدروکسیلی مورد نیاز برای واکنش با کربن کربونیلی کایرال نباشد، نیاز به انجام مرحله ۴ نیست.
مثال اول
چه کسی عبارت درشت مولکول ها را برای اولین بار به کار برد؟
پاسخ: «هرمان اشتاودینگر» (Herman Staudinger) اولین نفری بود که از این عبارت استفاده کرد.
مثال دوم
عبارت زیر را با واژه مناسب تکمیل کنید.
درشت مولکول ها از واحدهای تکرارشوندهای به نام ... ساخته شدهاند.
پاسخ: این واحدهای تکرارشونده، مونومر نامیده میشوند.
مثال سوم
درشت مولکولی که گلوکز را در بدن حیوانات ذخیره میکند، چه نام دارد؟
پاسخ: گلیکوژن درشت مولکولی است که گلوکز را در بدن حیوانات ذخیره میکند.
مثال چهارم
جال خالی را با واژه مناسب پر کنید.
... بسپاری متشکل از سه تکپار متفاوت است.
پاسخ: سهبسپار
مثال پنجم
تفاوت تکپار و پیشبسپار در چیست؟
پاسخ: پیشبسپارها از تکپارها پیچیدهتر هستند اما از بسپارها استحکام کمتری دارند.
مثال ششم
اتصال دو آمینواسید از چه طریقی است؟
پاسخ: اتصال بین گروه کربوکسیلیک اسیدی در آمینواسید اول و آمین آمینواسید دوم
مثال هفتم
به مولکولهایی که در خود هم گروه قطبی و هم گروه ناقطبی دارند، چه نام دارند؟
پاسخ: به این مولکولها آمفیپاتیک گفته میشود.
مثال هشتم
تمرین و آزمون
مثال نهم
تمرین و آزمون
مثال دهم
تمرین و آزمون
مثال یازدهم
تمرین و آزمون
مثال دوازدهم
تمرین و آزمون
مثال سیزدهم
تمرین و آزمون
مثال چهاردهم
تمرین و آزمون
مثال پانزدهم
تمرین و آزمون
مثال شانزدهم
تمرین و آزمون
مثال هفدهم
تمرین و آزمون
مثال هجدهم
تمرین و آزمون
مثال نوزدهم
تمرین و آزمون
مثال بیستم
تمرین و آزمون
مثال بیست و یکم
تمرین و آزمون
مثال بیست و دوم
تمرین و آزمون
مثال بیست و سوم
تمرین و آزمون
مثال بیست و چهارم
تمرین و آزمون
مثال بیست و پنجم
تمرین و آزمون
مثال بیست و ششم
تمرین و آزمون
مثال بیست و هفتم
تمرین و آزمون
مثال بیست و هشتم
تمرین و آزمون
مثال بیست و نهم
تمرین و آزمون
مثال سیام
تمرین و آزمون
مثال سی و یکم
تمرین و آزمون
مثال سی و دوم
تمرین و آزمون
مثال سی و سوم
تمرین و آزمون
مثال سی و چهارم
تمرین و آزمون
مثال سی و پنجم
تمرین و آزمون
مثال سی و ششم
تمرین و آزمون
مثال سی و هفتم
تمرین و آزمون
مثال سی و هشتم
تمرین و آزمون
مثال سی و نهم
تمرین و آزمون
مثال چهلم
تمرین و آزمون
مثال چهل و یکم
تمرین و آزمون
مثال چهل و دوم
تمرین و آزمون
مثال چهل و سوم
تمرین و آزمون
مثال چهل و چهارم
تمرین و آزمون
مثال چهل و پنجم
تمرین و آزمون
مثال چهل و ششم
تمرین و آزمون
مثال چهل و هفتم
تمرین و آزمون
مثال چهل و هشتم
تمرین و آزمون
مثال چهل و نهم
تمرین و آزمون
مثال پنجاهم
تمرین و آزمون
مثال پنجاه و یکم
تمرین و آزمون
مثال پنجاه و دوم
تمرین و آزمون
مثال پنجاه و سوم
تمرین و آزمون
مثال پنجاه و چهارم
تمرین و آزمون
مثال پنجاه و پنجم
تمرین و آزمون
مثال پنجاه و ششم
تمرین و آزمون
مثال پنجاه و هفتم
تمرین و آزمون
مثال پنجاه و هشتم
تمرین و آزمون
مثال پنجاه و نهم
تمرین و آزمون
مثال شصتم
تمرین و آزمون
مثال شصت و یکم
تمرین و آزمون
مثال شصت و دوم
تمرین و آزمون
مثال شصت و سوم
تمرین و آزمون
مثال شصت و چهارم
تمرین و آزمون
مثال شصت و پنجم
تمرین و آزمون
مثال شصت و ششم
تمرین و آزمون
مثال شصت و هفتم
تمرین و آزمون
مثال شصت و هشتم
تمرین و آزمون
مثال شصت و نهم
تمرین و آزمون
مثال هفتادم
تمرین و آزمون
مثال هفتاد و یکم
تمرین و آزمون
مثال هفتاد و دوم
تمرین و آزمون
مثال هفتاد و سوم
تمرین و آزمون
مثال هفتاد و چهارم
تمرین و آزمون
مثال هفتاد و پنجم
تمرین و آزمون
مثال هفتاد و ششم
تمرین و آزمون
مثال هفتاد و هفتم
تمرین و آزمون
مثال هفتاد و هشتم
تمرین و آزمون
مثال هفتاد و نهم
تمرین و آزمون
مثال هشتادم
تمرین و آزمون
مثال هشتاد و یکم
تمرین و آزمون
مثال هشتاد و دوم
تمرین و آزمون
مثال هشتاد و سوم
تمرین و آزمون
مثال هشتاد و چهارم
تمرین و آزمون
مثال هشتاد و پنجم
تمرین و آزمون
مثال هشتاد و ششم
تمرین و آزمون
مثال هشتاد و هفتم
تمرین و آزمون
مثال هشتاد و هشتم
تمرین و آزمون
مثال هشتاد و نهم
تمرین و آزمون
مثال نودم
تمرین و آزمون
سوالات متداول
حال که میدانیم درشت مولکولها چه ساختاری دارند، تصمیم داریم به برخی از مهمترین و پرتکرارترین سوالهای پیرامون این موضوع، پاسخ دهیم.

چهار گروه درشت مولکول های زیستی را نام ببرید ؟
درشت مولکولهای زیستی شامل کربوهیدرات، پروتئین، لیپید و نوکلئیک اسید هستند.
اهمیت درشت مولکول ها در چیست ؟
بسیاری از مواد غذایی مورد استفاده در رژیم غذایی انسان از درشت مولکولها ساخته شده است. این مواد به بدن کمک میکنند تا عملکرد خود را بهخوبی انجام دهد.
پروتئین چه درشت مولکولی است ؟
پروتئین درشت مولکولی دارای ۴ مرحله ساختار متنوع است که از توالی آمینواسیدهای مختلفی ساخته شدهاند.
کدام درشت مولکول در خود انرژی ذخیره می کند ؟
کربوهیدارتها درشت مولکولهایی هستند که انرژی بدن را تامین میکنند. از جمله میتوان به گلیکوژن اشاره کرد.
چرا درشت مولکول DNA از RNA پایدارتر است ؟
درحالی که DNA دارای دئوکسی ریبوز است، RNA ریبوز دارد. حضور الکل نوع دوم روی حلقه پنتوزی باعث کاهش پایداری RNA میشود.
کدام باز نیتروژنی است که در RNA موجود است اما در DNA خیر؟
اوراسیل باز نیتروژنی است که تنها در درشت مولکول RNA وجود دارد.
آیا لیپیدها انرژی را ذخیره می کنند ؟
بله، لیپیدها منبعی برای ذخیره انرژی در بدن انسان هستند.
آیا لیپیدها در آب حل می شوند ؟
خیر، لیپیدها مولکولهایی هستند که در حلال غیرقطبی حل میشوند و در آب که قطبی است، حل نمیشوند.
از بین دو درشت مولکول DNA و RNA کدام واکنش پذیرتر هستند ؟
از آنجا که RNA دارای قند ریبوز است، از DNA واکنشپذیری بیشتری دارد، بهصورتی که در محیط بازی پایدار نیست.
کدام باز نیتروژنی در DNA وجود دارد که در RNA دیده نمیشود؟
باز نیتروژنی تیمین در DNA وجود دارد و در RNA خیر.
RNA و DNA چه شباهت هایی دارند ؟
این دو درشت مولکول در ۳ مورد با یکدیگر شباهت دارند.
- از واحدهای تکپاری به نام نوکلئوتید ساخته شدهاند.
- هر دو دارای ۴ باز نیتروژنی هستند.
- هر دو دارای اسکلت فسفاتی هستند که به باز ساختار متصل شدهاند.
جمع بندی
در این مطلب در مورد درشت مولکول ها و انواع آنها صحبت کردیم. درشت مولکولهای زیستی نقش بسیار مهمی در حیات روی کره زمین دارند و شامل: پروتئين، نوکلئیک اسید، لیپید و کربوهیدراتها هستند. هر کدام از این درشت مولکول ها خانواده بزرگی را تشکیل میدهند که بهتفضیل در مورد نقش هر کدام از آنها صحبت شده است.
آزمون درشت مولکول ها
۱. در تعریف علمی درشتمولکول، چه ویژگی ساختاری آن را از مولکولهای کوچکتر متمایز میکند؟
توانایی واکنش با آب و حل شدن آسان در آن
تعداد بالای اتمها در ساختار و زنجیره بسیار طولانی
رفتار الکتریکی متفاوت نسبت به یونها در محیط آبی
حضور همزمان کربن و نیتروژن در ترکیب شیمیایی
ویژگی اصلی «تعداد بالای اتمها در ساختار و زنجیره بسیار طولانی» موجب تمایز درشتمولکولها از مولکولهای کوچکتر میشود. این ترکیبات معمولا دارای بیش از ده هزار اتم و ساختار زنجیرهای بسیار بزرگ هستند که از اتصال پیدرپی مونومرها به دست میآیند. سایر ویژگیها مانند وجود عناصر خاص، قابلیت حل شدن آسان در آب یا رفتار الکتریکی، به طور معمول بین انواع مولکولها مشترک یا محدود به گروههای خاصی است و ویژگی تعریفکننده همه درشتمولکولها نیستند.
۲. واژههای Poly و Mer در اصطلاح بسپار (Polymer) چگونه به شناخت ساختار این نوع درشتمولکولها کمک میکنند؟
Poly به معنی چند و Mer به معنی واحد، یعنی بسپار از تکرار چند واحد ساخته میشود.
Poly به معنی پیوند و Mer به معنی مولکول، یعنی بسپار فقط از پیوند مولکولها ایجاد میشود.
Poly نشاندهنده ساختار حلقوی و Mer نمایانگر قند، یعنی بسپار فقط از قندها ساخته میشود.
Poly به معنی زنجیره و Mer به معنی آمینواسید، یعنی بسپارها زنجیرهای از آمینواسید هستند.
عبارت «Poly به معنی چند» و «Mer به معنی واحد» ریشه یونانی دارند و بیان میکنند که ساختار بسپار (Polymer) از تکرار چندین واحد سازنده کوچکتر بهنام مونومر به وجود میآید.
۳. تفاوت اصلی ساختاری مونومر و پلیمر چیست و هر کدام چه نقشی در شکلگیری درشتمولکولها دارند؟
هر دو ساختار مشابه دارند اما مونومر بیشتر در سنتز پروتئین استفاده میشود.
پلیمر و مونومر فقط در لیپیدها یافت میشوند و نقش مشابهی دارند.
پلیمر ساختار کوچکتری دارد و مونومر ترکیب بزرگ و پیچیده است.
مونومر واحد سازنده تکرارشونده است و پلیمر زنجیرهای از این واحدهاست.
مونومر یک مولکول ساده است که به صورت تکرارشونده با یکدیگر واکنش میدهند و تشکیل پلیمر میدهند. این پلیمرها در واقع ساختار اصلی درشتمولکولهای زیستی نظیر کربوهیدراتها، پروتئینها و اسیدهای نوکلئیک را میسازند.
۴. کدام مورد شامل چهار گروه اصلی درشتمولکولهای زیستی است؟
کربوهیدرات، پروتئین، لیپید، اسید نوکلئیک
کربوهیدرات، ویتامین، لیپید، مواد معدنی
اسید نوکلئیک، هورمون، پروتئین، ویتامین
پروتئین، ویتامین، آنزیم، اسید نوکلئیک
ترکیب «کربوهیدرات، پروتئین، لیپید و اسید نوکلئیک» همان چهار گروه اصلی شناختهشده درشتمولکولهای زیستی است که نقشهای ساختاری و عملکردی کلیدی در سلول دارند.
۵. کربوهیدراتها عمدتا از چه عناصر شیمیایی ساخته شدهاند و این ترکیب چه تاثیری بر ساختار آنها دارد؟
کربن، اکسیژن و هیدروژن. باعث تکرار واحدهای ساده ساختاری میشود.
کربن، منیزیم و کلر. استحکام ساختار حلقوی آنها را تعیین میکند.
هیدروژن، گوگرد و سدیم. موجب انعطاف ساختاری در آنها میشود.
کربن، نیتروژن و فسفر. سبب خاصیت اسیدی آنها است.
کربوهیدراتها عمدتا از عناصر کربن، اکسیژن و هیدروژن تشکیل شدهاند. این ترکیب سهگانه باعث ایجاد ساختارهای تکرارشونده در آنها میشود که واحدهای سادهای مثل مونوساکاریدها میتوانند به صورت پلیساکارید زنجیره شوند. سایر گزینهها با عناصر نادرست و ویژگیهای نادرست ساختاری، مرتبط با کربوهیدراتها نیستند.
۶. کربوهیدراتها در زیستشناسی به چند گروه اصلی تقسیم میشوند و ویژگی کلیدی هر گروه چیست؟
سه گروه: مونوساکارید، دیساکارید و پلیساکارید؛ بر اساس تعداد واحد قندی
چهار گروه: نشاسته، سلولز، گلیکوژن و فروکتوز؛ با پایه ساختاری متفاوت
دو گروه: احیاکننده و غیر احیاکننده؛ بر اساس قابلیت واکنش شیمیایی
سه گروه: کربندار، هیدروژندار و اکسیژندار؛ براساس عنصر سازنده
کربوهیدراتها بر اساس شمار واحدهای قندی به سه گروه مونوساکارید، دیساکارید و پلیساکارید دستهبندی میشوند که شاخص آنها اندازه و تعداد تکرار واحدهای قند است.
۷. کدام مورد به بهترین شکل نقش زیستی کربوهیدراتها را در فرایندهای فتوسنتز و متابولیسم انرژی بدن توضیح میدهد؟
کربوهیدراتها فقط به عنوان ساختار سلولی در بدن موجودات زنده کاربرد دارند.
کربوهیدراتها صرفا در ساخت دیواره سلولهای جانوری شرکت دارند.
کربوهیدراتها منبع اصلی انرژی بوده و از طریق فتوسنتز در گیاهان تولید میشوند.
کربوهیدراتها فقط به عنوان انتقالدهنده پیامهای شیمیایی عمل میکنند.
عبارت «کربوهیدراتها منبع اصلی انرژی بوده و از طریق فتوسنتز در گیاهان تولید میشوند» صحیح است، زیرا کربوهیدراتها در گیاهان طی واکنش فتوسنتز ساخته میشوند و سپس در بدن انسان و جانوران، به وسیله متابولیسم به انرژی تبدیل میگردند. نقش ساختاری نیز ممکن است وجود داشته باشد، اما محدود به کربوهیدراتها نیست.
۸. در مولکول گلوکز، مراکز کایرال چه نقشی در ایجاد ایزومرهای مختلف ایفا میکنند؟
مراکز کایرال فقط بر ویژگی آبدوست بودن گلوکز اثر میگذارند.
مراکز کایرال فقط نشاندهنده گروههای عامل اکسیژن هستند.
ایزومرهای گلوکز به دلیل تغییر موقعیت پیوندهای گلیکوزیدی ایجاد میشوند.
وجود مراکز کایرال باعث تولید فرمهای نوری D و L در گلوکز میشود.
در گلوکز، وجود مراکز کایرال موجب شکلگیری دو فرم نوری D و L میشود، زیرا ترتیب متفاوت گروهها اطراف این مراکز ساختارها را متمایز میکند. گزینه «مراکز کایرال فقط نشاندهنده گروههای عامل اکسیژن هستند» صحیح نیست زیرا مراکز کایرال علاوه بر اکسیژن، به چهار گروه متفاوت متصلاند.
۹. تفاوت اصلی ساختاری فرم آلفا و بتا گلوکز در حالت حلقوی چیست و این تفاوت چه اثری بر عملکرد زیستی آنها دارد؟
آلفا و بتا تفاوتی در موقعیت اتم اکسیژن ندارند اما قابلیت انحلالپذیری متفاوت دارند.
جهت قرارگیری گروه هیدروکسیل روی کربن شماره یک در آلفا و بتا متفاوت است و این تفاوت نوع پیوند پلیساکاریدی را تعیین میکند.
تعداد اتمهای کربن در حلقه آلفا کمتر از بتا است، بنابراین فقط یکی از این فرمها میتواند انرژی را ذخیره کند.
در فرم آلفا فقط ایزومر D تشکیل میشود در حالیکه فرم بتا به هر دو صورت D و L وجود دارد.
تفاوت ساختاری میان فرم آلفا و بتا گلوکز در جهت قرارگیری گروه هیدروکسیل روی اتم کربن شماره یک است؛ در آلفا و بتا این گروه به بالا یا پایین حلقه قرار میگیرد که نتیجه آن نوع پیوند قابل تشکیل در پلیساکاریدها مانند نشاسته یا سلولز است. این موضوع باعث میشود ترکیبات حاصل از هر فرم خواص شیمیایی و زیستی مجزایی پیدا کنند.
۱۰. فرایند تولید دیساکارید و پلیساکارید توسط پیوند گلیکوزیدی بین مولکولهای ساکاریدی چگونه انجام میشود؟
اتصال مونوساکاریدها از طریق پیوندهای گلیکوزیدی زنجیرهای ایجاد میکند.
تعویض گروه فسفات مونوساکاریدها باعث یکیشدن آنها میشود.
افزودن گروه آمینه به هر مونوساکارید، ترکیب بزرگ تولید میکند.
شکستن پلیمرهای موجود و اتصال قطعات به هم مولکول جدید میسازد.
در ساخت دیساکارید و پلیساکارید، مونوساکاریدها طی واکنش بسپارش با تشکیل پیوند گلیکوزیدی به یکدیگر متصل میشوند و ساختارهای بلند و زنجیرهای ایجاد میکنند.
۱۱. از نظر ساختار مولکولی و کارکرد زیستی، چه تفاوتی میان سلولز و نشاسته وجود دارد؟
سلولز با واحدهای گلوکز با پیوند هیدروژنی کنار هم قرار میگیرد و نشاسته به فرم لیپید است.
هر دو با پیوندهای β گلیکوزیدی ساخته میشوند ولی سلولز انرژی ذخیره میکند و نشاسته ساختاری است.
سلولز با پیوندهای β گلیکوزیدی ساخته میشود و نقش ساختاری در دیواره سلول گیاهی دارد، نشاسته با پیوندهای α گلیکوزیدی ساخته میشود و ذخیره انرژی است.
سلولز و نشاسته هر دو ذخیره انرژی هستند اما ساختار گلوکز متفاوت دارند.
طبق دادههای علمی موجود، پیوند بین گلوکزها در سلولز نوع β است و این پلیساکارید در دیواره سلول گیاهی برای استحکام ساختاری بهکار میرود. در حالی که نشاسته حاصل پیوند α گلیکوزیدی میان گلوکزهاست و نقش اصلی آن ذخیره انرژی در گیاهان است.
۱۲. کدام مورد تفاوت مهم دیساکارید کاهنده و غیرکاهنده را توضیح میدهد؟
دیساکارید غیرکاهنده توانایی جذب آب بیشتری نسبت به کاهنده دارد.
دیساکارید کاهنده دارای پیوندی است که یک گروه آلدهیدی فعال باقی میماند.
دیساکارید کاهنده همیشه شامل یک گروه آمینی است.
دیساکارید غیرکاهنده قابلیت بیرنگ شدن محلول را ندارد.
در دیساکارید کاهنده، پیوند بین دو مونوساکارید به گونهای است که حداقل یک گروه آلدهیدی یا کتونی آزاد میماند و قابلیت شرکت در واکنشهای شیمیایی خاص مانند اکسایش را دارد. این ویژگی باعث اهمیت شیمیایی و واکنشپذیری بیشتر این ترکیبات میشود.
۱۳. در واکنشهای هیدرولیز و واسرشتگی (Denaturation) پروتئینها، افزایش دما یا تغییر pH چه اثری بر ساختار و عملکرد پروتئین دارد؟
افزایش دما یا تغییر pH باعث تغییر ساختار سهبعدی و کاهش عملکرد پروتئین میشود.
بر اثر افزایش دما یا تغییر pH، تعداد آمینواسیدهای پروتئین افزایش مییابد.
افزایش دما یا تغییر pH فقط پیوند پپتیدی را تقویت میکند و ساختار ثابت میماند.
تحت افزایش دما یا تغییر pH، پروتئین به یک مولکول لیپید تبدیل میشود.
تغییرات دما یا pH میتواند ساختار سهبعدی پروتئین را مختل کند که این فرآیند دناتوره شدن نام دارد و عملکرد زیستی پروتئین کاهش مییابد.
۱۴. ویژگی آمفیپاتیک فسفولیپیدها چه نقشی در تشکیل ساختار دو لایه غشای سلولی دارد؟
تشکیل لایه منفرد فسفولیپیدی و جلوگیری از برهمکنش با آب را ممکن میسازد.
اجزای آبدوست و آبگریز فسفولیپیدها باعث آرایش دو لایه با دم به داخل و سر به خارج میشوند.
باعث قرارگیری دمهای آبگریز به سمت خارج سلول میشود.
هیدروفیلی بودن کامل فسفولیپیدها باعث انتقال آزاد مواد از غشا میشود.
فسفولیپیدها دارای سر آبدوست و دم آبگریز هستند. این ویژگی آمفیپاتیک باعث میشود سرهای آبدوست به سمت محیط آبی درون و بیرون سلول قرار گیرند و دمهای آبگریز درون غشا و دور از آب به هم نزدیک شوند. در نتیجه، فسفولیپیدها بهصورت دو لایه منظم قرار میگیرند و ساختار غشای سلولی را شکل میدهند.
۱۵. از نظر ساختار و نقش زیستی، کدام مورد درباره تفاوت اسیدهای چرب اشباع و غیراشباع صحیح است؟
اسید چرب اشباع حاوی پیوند دوگانه و در دیواره سلولی نقش دارد.
اسید چرب اشباع فقط در گیاهان وجود دارد و آبگریز نیست.
اسید چرب غیراشباع تنها در حیوانات یافت میشود و ساختاری خطی دارد.
اسید چرب غیراشباع دارای یک یا چند پیوند دوگانه در زنجیره است.
اسیدهای چرب غیراشباع برخلاف اشباع، یک یا چند پیوند دوگانه دارند که روی ساختار و عملکرد زیستی آنها تاثیرگذار است.
۱۶. در کروماتوگرافی اندازه طردی، عامل اصلی جداسازی مولکولها چیست و این روش در علوم زیستی چگونه به کار میرود؟
جداسازی بر اساس بار الکتریکی مولکولها، برای خالصسازی پروتئینها
جداسازی براساس اندازه و جرم مولکولی، جهت تفکیک درشتمولکولهایی مانند پروتئین و DNA
جداسازی براساس پیوندهای شیمیایی، جهت بررسی واکنش آنزیمها
جداسازی بر اساس میزان حلالیت، برای شناسایی قندها و لیپیدها
در روش «جداسازی براساس اندازه و جرم مولکولی»، کروماتوگرافی اندازه طردی سبب میشود مولکولهای بزرگتر و کوچکتر بر اساس اندازه متفاوت خود جدا شوند و این تکنیک بهویژه برای تفکیک درشتمولکولهایی مانند پروتئینها، آنزیمها یا اسیدهای نوکلئیک (DNA و RNA) در علوم زیستی به کار میرود
۱۷. پروتئینهای مزدوج مانند گلیکوپروتئین در مقایسه با پروتئینهای ساده از نظر عملکرد زیستی چه تفاوتی دارند؟
پروتئینهای مزدوج ویژگی آبگریزی بیشتری نسبت به پروتئینهای ساده دارند.
پروتئینهای مزدوج قادرند با مولکولهای غیرپروتئینی مانند قند یا لیپید ترکیب شده و نقشهای متنوعتری ایفا کنند.
پروتئینهای مزدوج تنها از واحدهای آمینواسیدی ساخته میشوند و قابلیت انعطاف ساختاری بیشتری دارند.
پروتئینهای مزدوج فقط در ساختار سلولی نقش دارند و در واکنشهای زیستی شرکت نمیکنند.
پروتئینهای مزدوج مانند «گلیکوپروتئین» میتوانند با اجزای غیرپروتئینی از جمله قندها و لیپیدها ترکیب شوند، که این ترکیب باعث گسترش و تنوع عملکرد آنها در بدن میگردد. این گروه برخلاف پروتئینهای ساده که فقط از زنجیرههای آمینواسید تشکیل شدهاند، قابلیت انجام کارکردهایی مانند ارسال سیگنالهای زیستی یا حمل ملکولهای خاص را دارند.
۱۸. ساختار سهبعدی پروتئینها چه نقشی در عملکرد آنها در بدن دارد؟
شرایط اتصال مولکولهای دیگر به پروتئین را تعیین میکند.
باعث افزایش چربی بدن میشود.
امکان تشخیص قندها را فراهم میکند.
نقش ذخیرهسازی اطلاعات ژنتیکی را ایفا میکند.
ساختار سهبعدی پروتئینها باعث میشود که پروتئین بتواند به شکل اختصاصی با مولکولهای دیگر تعامل کند. این ساختار ویژه محلهای اتصال یا برهمکنش را بوجود میآورد و بر عملکرد زیستی پروتئین تاثیر مستقیم میگذارد.

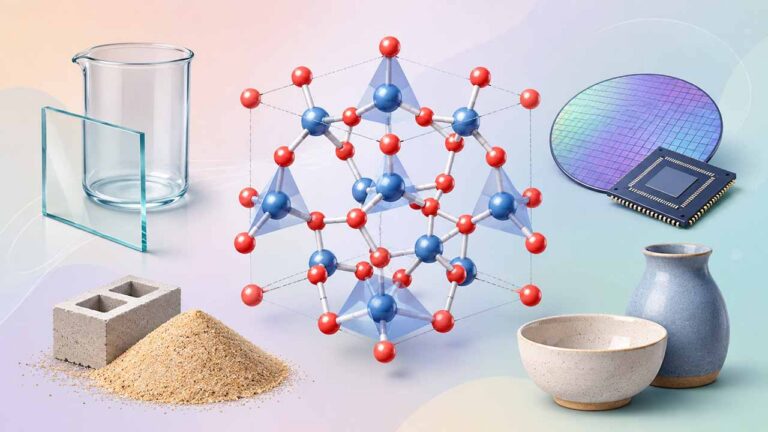





ممنونم از مطلب بسیار کاربردی و مفیدتان.
از مطلبتان اینطور استنباط میشود که یک درشتمولکول همیشه یک بسپار است، اما اگر اشنباه نکنم درشتمولکولهایی داریم که بسپار نیستند مثل لیپیدها و البته گلیکولیپیدها! اینطور نیست؟
با سلام؛
از بازخورد شما بسیار سپاسگزاریم. در پلیمرها، واحدهای تکرارشونده نیز داریم که با درشتمولکول متفاوت است. برای بهبود خوانایی متن، تغییرات کوچکی ایجاد شد.
با تشکر از همراهی شما با مجله فرادرس