



سلول عصبی چیست؟ – به زبان ساده + کار و وظیفه
بدن مهرهداران از تقسیم یک سلول به تعداد زیادی سلولهای دیگر تشکیل میشود. در مراحل اولیه رشد جنین، شکل و وظیفه این سلولها تغییر میکند و هر دسته سلولی برای انجام فعالیت مشخصی از سلول تمایز مییابد. به همین دلیل نه تنها شکل بلکه فعالیت کلیه با معده تفاوت دارد. سلول عصبی یا نورون یکی از سلولهای تمایزیافته مهرهداران ازجمله انسان است که شکل کاملا متفاوت و اندامکهای مشابه دیگر سلولهای بدن دارد. شکل این سلولها شبیه درختان است و از تعداد زیادی شاخه به نام دندریت،تنه جسم سلولی و آکسون، و ریشههایی به نام شاخههای آکسونی تشکیل میشود. از کنار هم قرار گرفتن این سلولها به همراه سلولهای غیرعصبی، بافت عصبی تشکیل میشود. بافت عصبی را میتوان به دو بخش مرکزی و محیطی تقسیمبندی کرد. بخش مرکزی شامل مغز و نخاع، و بخش محیطی شامل سلولهای عصبی انتقال پیام است. در این مطلب توضیح میدهیم ویژگیهای ظاهری متفاوت سلول عصبی چیست .
- یاد میگیرید که ساختار نورون و نقش اجزای آن چگونه است.
- میآموزید نورونها چگونه پیام عصبی را تولید و منتقل میکنند.
- خواهید آموخت سیستم عصبی چگونه بخشها و عملکردهای متنوع بدن را کنترل میکند.
- طبقهبندی انواع نورونها و تفاوت کارکردیشان را یاد خواهید گرفت.
- عملکرد گیرندههای حسی و نقششان در انتقال پیام را میآموزید.
- با سلولهای گلیا و نقش حمایتیشان در سیستم عصبی آشنا میشوید.
- تأثیر انتقالدهندههای عصبی در سیناپسها را بررسی خواهید کرد.
- خواهید توانست نقش جامع نورونها در هماهنگی بدن را توصیف کنید.




نورونها بهوسیله مولکولهای شیمیایی کوچک با هم ارتباط برقرار میکنند. در غشای دندریتهای هر نورون گیرندهای وجود دارد که به مولکولهای انتقال پیام متصل میشود و نفوذپذیری غشا به یونها را تغییر میدهد. پیام عصبی، جریان الکتریکی ایجاد شده از افزایش ورود یونها به سیتوپلاسم نورون است. این جریان از اندامهای مختلف ازجمله پوست انگشتان دست، سلولهای ماهیچهای لوله گوارش، ماهیچههای دست و چشمها به بخشهای مختلف مغز منتقل میشود. مغز پیام عصبی را پردازش و دستور مناسب آن را صادر میکند. در نتیجه فعالیتهای بدن تنظیم میشود. در این مطلب از مجله فرادرس توضیح میدهیم انواع و تحریک پذیری سلول عصبی چیست و این سلولها چه وظیفهای در بدن دارند.
سلول عصبی چیست؟
سلول عصبی یا نورون، سلولهای تشکیلدهنده سیستم عصبی مهرهداران هستند. این سلولها از جسم سلولی، آکسون و دندریتها تشکیل میشود. جسم سلولی بخشی است که بیشترین اندامکها و سیتوپلاسم را به خود اختصاص داده و در بیشتر نورونها بین دندریت و آکسون قرار دارد. هسته سلول، شبکه اندوپلاسمی صاف و زبر، جسم گلژی و لیزوزوم نورونها در جسم سلولی قرار دارد. دندریتها زوائد سیتوپلاسمی کوتاه شبیه به شاخههای درخت هستند که از جسم سلولی خارج میشود. در سیتوپلاسم این زوائد پروتئینهای اسکلت سلولی وجود دارد. زوائد سیتوپلاسمی کوچکتری به نام خار در دندریتها وجود دارد که محل برقراری ارتباط دو سلول عصبی است.
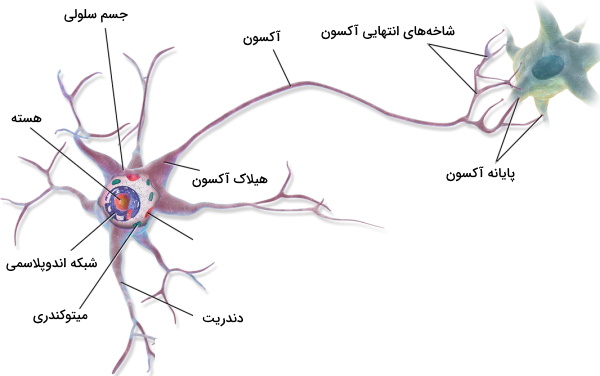
آکسون رشته سیتوپلاسمی بلندی است که از جسم سلولی خارج میشود. این رشته بهوسیله هیلاک به جسم سلولی متصل میشود. انتهای آکسون (بخش دور از جسم سلولی) به شاخههایی تقسیم میشود. قطر هر شاخه در انتها افزایش پیدا میکند و پایانه عصبی را تشکیل میدهد. پایانه عصبی هر آکسون با دندریت، جسم سلولی یا آکسون سلول عصبی دیگر ارتباط برقرار میکند. در هر رشته آکسون مجموعهای از پروتئینهای اسکلت سلولی به نام میکروتوبول، داینئین و کاینزین وجود دارد که به انتقال مواد از جسم سلولی به آکسون و آکسون به جسم سلولی کمک میکند. آکسون بسیاری از نورونهای عصبی بهوسیله لایه لیپیدی به نام غلاف میلین پوشیده شده است.
سیستم عصبی چیست؟
سیستم عصبی مهمترین سیستم کنترل، تنظیم و هماهنگی اندامها و دستگاههای بدن است که به کمک سیستم اندوکرین به حفظ هموستازی بدن کمک میکند. این سیستم از نورونها و سلولهای پشتیبان، بافت پیوندی و رگهای خونی تشکیل شده است. سیستم عصبی پیام بخشهای مختلف بدن و محیط اطراف را بهوسیله نورونهای حسی دریافت میکند و پس از پردازش در مغز پاسخ مناسب را صادر میکند. این سیستم را میتوان به دو بخش مرکزی و محیطی تقسیم کرد. سیستم عصبی مرکزی از مغز و نخاع تشکیل میشود. مغز مرکز کنترل اصلی سیستم عصبی است که بهوسیله نورونهای نخاع با سیستم عصبی محیطی ارتباط برقرار میکند. سیستم عصبی محیطی مجموعهای از نورونهای حسی و حرکتی است که پیام عصبی را بین سیستم عصبی مرکزی و اندامهای مختلف بدن جابهجا میکند. مغز از سه بخش اصلی مخ، مخچه و ساقه مغز تشکیل شده است.
- مخ: مخ بزرگترین بخش مغز است که در ادغام پیامهای حسی، کنترل حرکات ارادی بدن و فعالیتهای پییچدهتر مثل تفکر انتزاعی و تکلم نقش دارد. به لایه خارجی این بخش از مغز، قشر مخ گفته میشود. قشر مخ از قسمتهای بدون میلین نورون ها با شیارها و برآمدگیهای زیادی تشکیل شده است. قسمتهای میلیندار نورونها در بخشهای داخلی مخ قرار گرفته است. مخ بهوسیله یک شیار عمیق به نیمکره راست و چپ تقسیم میشود. این دو نیمکره بهوسیله دستهای از سلولهای عصبی میلیندار به نام جسم پسنهای با هم ارتباط برقرار میکنند. هر نیمکره حرکت سمت مخالف بدن را کنترل میکند. هر نیمکره مخ به چهار بخش یا لوب جلویی، آهیانه، گیجگاهی و پسسری تقسیم میشود و سلولهای عصبی هر بخش فعالیتهای متفاوتی را کنترل میکنند. برای مثال پیامهای عصبی چشم در لوب پسسری و پیامهای عصبی گوش در لوب گیجگاهی پردازش میشود. در بخشهای داخلی مخ حفرههایی به نام بطن وجود دارد که از مایع مغزی-نخاعی پر شده است.
- مخچه: مخچه مثل مخ بهوسیله دستهای از سلولهای عصبی میلیندار به نام کرمینه به دو نیمکره چپ و راست تقسیم میشود. قشر مخچه از نورونهای بدون میلین و بخشهای داخلی آن از قسمتهای میلندار نورون تشکیل شده است. این بخش مغز در تنظیم تعادل، کشش طبیعی ماهیچهها و هماهنگی حرکات ارادی اندامها را کنترل میکند.
- ساقه مغز: ساقه مغز مجموعهای از سلولهای عصبی است که بین مخ، مخچه و نخاع ارتباط برقرار میکند و از سه بخش اصلی مغز میانی، پل مغزی و بصل النخاع تشکیل شده است.
انواع سلول عصبی چیست؟
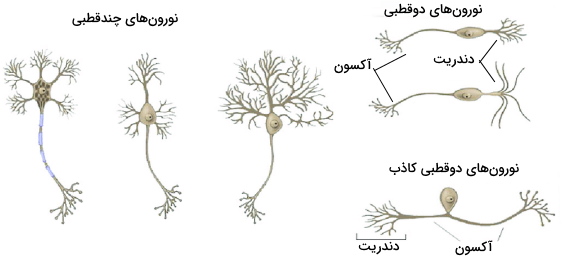
حالا که متوجه شدیم سلول عصبی چیست ، میتوانیم انواع آن را بررسی کنیم. سلولهای عصبی را میتوان بر اساس شکل و عملکرد به انواع مختلف تقسیم کرد. این سلولها بر اساس شکل به انواع چندقطبی، دوقطبی، دوقطبی کاذب و تکقطبی تقسیم میشود. در نورونهای چندقطبی یک رشته آکسون از یک طرف جسم سلولی و تعداد زیادی دندریت از طرف دیگر جسم سلولی خارج میشود. در نورونهای دوقطبی دو رشته بلند از دو طرف جسم سلولی خارج میشود. یکی از این رشتهها دندریت و دیگری آکسون است. رشته دندریتی به شاخههای زیادی تقسیم میشود.
در نورونهای دوقطبی کاذب، رشته سیتوپلاسمی کوتاهی از جسم سلولی خارج و به دو بخش اصلی دندریت و آکسون تقسیم میشود. در نورونهای تکقطبی یک رشته سیتوپلاسمی بلند از جسم سلولی خارج میشود و دندریت و آکسون را تشکیل میدهد. جسم سلولی این نورونها در یکی از دو انتهای سلول، دندریتها وسط و آکسون در انتهای دیگر قرار دارد. این نورونها بخشهای مختلف سیستم عصبی را میسازند.
بر اساس عملکرد، نورونها را به انواع حسی، حرکتی و بینابینی یا رابط تقسیم میکنیم. نورونهای حسی اطلاعات مربوط به اتفاقات بخشهای مختلف داخل بدن و محیط خارج از بدن را به بافت عصبی در مغز و نخاع منتقل میکند. به این سلولهای عصبی نورونهای آوران نیز گفته میشود و از انواع نورونهای دوقطبی کاذب هستند. برای مثال اگر گلوله برفی را در دست بگیرید، نورونهای حسی در پوست انگشتان دست تغییر دما را به مغز منتقل و شما سرما را احساس میکنید. نورونهای حرکتی دستور مغز را به اندامها دیگر منتقل میکنند. به این سلولهای عصبی نورون وابران نیز گفته میشود و از انواع نورونهای چندقطبی هستند. برای مثال پیام عصبی مغز سبب انقباض ماهیچههای پا و فرار کردن از خرس میشود. نورونهای بینابینی در مغز و نخاع پیام عصبی را از یک نورون دریافت (نورون حسی یا بینابینی) و به نورون دیگر (نورون حرکتی یا بینابینی) منتقل میکنند. در بافت تعداد نورونهای بینابینی از نورونهای حسی و حرکتی بیشتر است. این نورونها نقش مهمی در پردازش اطلاعات ورودی از بخشهای مختلف بدن و ایجاد پیام واحد در مغز دارند.
انواع نورون های حسی
نورونهای حسی را میتوان بر اساس بافتی که پیام عصبی آن را دریافت میکنند به انواع نورونهای حسی محیطی و احشایی تقسیم کرد. نورونهای حسی محیطی پیام عصبی پوست، بینی، گوش، چشم و زبان، اما نورونهای حسی احشایی پیام عصبی اندامهای داخلی ازجمله اندامهای حفره شکمی را به مغز منتقل میکنند. محرک نورونهای حسی محیطی تغییر دما و فشار پوست، مولکولهای شیمیایی بو، حرکت ایجاد شده از امواج صوتی، نور و مولکولهای شیمیایی مزه، اما محرک نورونهای حسی احشایی تغییرات فشار خون، درد، کشیدگی دیواره لوله گوارش، درد و تغییر غلظت مواد شیمیایی است. انتهای این نورونها گیرنده حسی است یا با یک گیرنده حسی در ارتباط است. گیرندههای حسی را بر اساس ساختار و نوع محرک به گروههای متفاوت تقسیم میشوند.
- انواع گیرنده حسی بر اساس ساختار: گیرندههای حسی بر اساس ساختار به سه گروه انتهای دندریتی نورون، انتهای دندریتی در کپسولی از بافت پیوندی و سلولها عصبی یا غیرعصبی تخصصیافته هستند. گیرندههای دما و درد پوست انتهای دندریت بدون کپسول، گیرنده فشار پوست انتهای دندریتی همراه کپسول، گیرندههای نوری چشم و بویایی نورونهای تمایزیافته و گیرندههای چشایی سلولهای بافت پوششی تمایزیافته هستند.
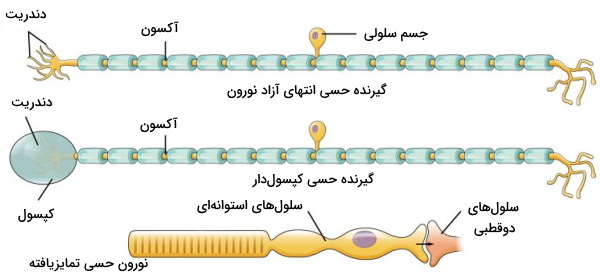
- انواع گیرنده حسی بر اساس محرک: محرکهای گیرندههای شیمیایی تغییرات غلظت یونها یا درشتمولکولهای محیط، تغییر ویژگیهای فیزیکی محیط و پرتوهای الکترومغناطیسی نور مرئی است. بر اساس محرک گیرندههای حسی را به انواع مکانیکی، شیمیایی، الکترومغناطیسی، فشاری و دمایی تقسیم میکنیم.
گیرنده های حس لامسه
حس لامسه مجموعهای از حسها است که در اثر لمس، درد و تغییر موقعیت ایجاد میشود. گیرندههای دما انواعی از گیرندههای حس لامسه هستند که در پوست و بخشهای داخلی بدن قرار دارند. بعضی از این گیرندهها با افزایش و بعضی از آنها با کاهش دما تحریک میشود. افزایش محرکهای مکانیکی، شیمیایی و تغییر دما ممکن است با تحریک احساس درد همراه باشد. مولکولهای شیمیایی آزاد شده از بافتهای آسیبدیده گیرندههای درد را فعال میکند. این گیرندهها دندریت آزاد یا کپسولدار هستند که تعداد آنها در پوست بیشتر از سایر بخشهای بدن است. دیسکهای مرکل، کپسولهای مایسنر، انتهای رافینی و اجسام پاچینی گیرندههای مکانیکی حس لامسه در پوست انسان هستند.
- دیسکهای مرکل: این گیرندهها در بخشهای سطحی، نزدیک خارجیترین لایه پوست (اپیدرم) دارای مو و بدون مو (کف پا، کف دست، لبها و انگشتان) قرار دارند. این گیرندهها انتهای دندریتی بدون کپسول هستند که در اثر فشارهای آرام روی پوست تحریک میشود. این گیرندهها تغییرات فشار را در بخش کوچکی از محیط اطراف خود شناسایی میکنند.
- کپسولهای مایسنر: کپسولهای مایسنر در بخشهای پایینتر از دیسکهای مرکل قرار دارند. این گیرندهها دندریتهای کپسولدار نورون حسی هستند که در پوست بدون موی انگشتان و پلک قرار دارند. مایع درون کپسول این گیرندهها علاوه بر لمس و فشار بر اثر لرزشهای خفیف نیز حرکت میکند و پیام عصبی ایجاد میشود.
- انتهای رافینی: انتهای رافینی، دندریتهای نورون حسی کپسولدار هستند که در بخشهای عمقیتر پوست بدون مو و مودار قرار دارند. این گیرندهها در اثر کشش پوست، تغییر شکل مفصلها و افزایش دما تحریک میشوند. به همین دلیل نقش مهمی در گرفتن اجسام و کنترل حرکت انگشتان دارند.
- اجسام پاچینی: ساختار اجسام پاچینی شبیه کپسولهای مایسنر است. این گیرندهها در بخشهای عمقی پوست مودار و بدون مو، پوشش اطراف استخوان (پریاُستیوم)، کپسول مفصلی، اندامهای حفره شکمی، پستان و اندامهای تولید مثلی قرار دارند. این گیرندهها بر اثر فشار عمقی لحظه و لرزشهای شدید تحریک میشوند. تعداد اجسام پاچینی و انتهای رافینی در پوست کمتر از دیسکهای مرکل و کپسول مایسنر است.
دوک ماهیچهای و اجسام گلژی تاندون، انواع دیگری از گیرندههای حس لامسه هستند که در تشخیص موقعیت اندامهای حرکتی در فضا کمک میکنند. دوک ماهیچهای از دو نوع فیبر ماهیچهای تشکیل شده که با نورونهای حسی سیناپس میدهند و در تشخیص کشیدگی و طول ماهیچهها کمک میکنند. اجسام گلژی تاندون از رشتههای تاندونی تشکیل شده که در کپسولی از بافت پیوندی قرار دارند و با دندریت نورونهای حسی سیناپس تشکیل میدهند. این گیرندهها با تغییر نیروی انقباضی ماهیچه تحریک میشوند.
بارورسپتورها
بارورسپتورها انتهای دندریتی نورونهای حسی و بدون کپسول هستند. این گیرندهها در سرخرگهای آئورت و کاروتید، دهلیزها، بطنها و رگهای ریه قرار دارند و در اثر افزایش یا کاهش فشار و حجم خون تحریک میشوند. وظیفه بارورسپتورهای ثابت نگهداشتن فشار خون است. این گیرندهها را میتوان به دو گروه بارورسپتورهای فشار بالا و پایین تقسیمبندی کرد.
- بارورسپتورهای فشار بالا: این گیرندهها تغییرات کوچک فشاره خون در شرایط فیزیولوژیک بدن را تنظیم میکند. اما در شرایط افزایش فشار خون بالا برای مدت طولانی (بیش از ۲ تا ۳ روز)، با شرایط سازگار میشوند و آستانه تحریک آنها افزایش مییابد. کشش غشای نورون به دلیل افزایش فشار خون با ایجاد پتانسیل عمل، انتقال پیام عصبی به مراکز کنترل فشار خون در مغز، کاهش ضربان قلب و در نهایت کاهش فشار خون همراه است. کاهش فشار خون تحریک این سلولها را کاهش و ضربان قلب را افزایش میدهد. در نتیجه فشار خون افزایش مییابد.
- بارورسپتورهای فشار پایین: این گیرندهها در دیواره دهلیزها و سیاهرگهای بزرگ بدن قرار دارند و به تنظیم حجم خون کمک میکنند. تحریک این گیرندهها علاوه بر تغییرات سیستم عصبی، ترشح هورمونهای تحریککننده بازجذب نمکها از کلیه را افزایش میدهد.
گیرنده نوری
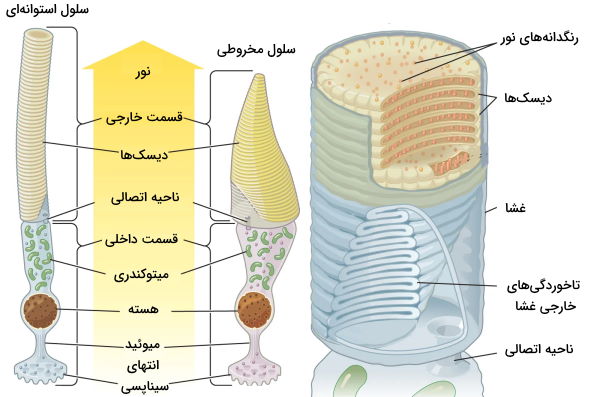
گیرندههای نوری سلولهای عصبی تمایزیافتهای هستند که در شبکیه چشم قرار دارند و نور ورودی به چشم را به پیام عصبی تبدیل میکنند. سلولهای استوانهای و عصبی دو گیرنده نوری چشم انسان هستند که شکل و عملکرد متفاوتی دارند. ناحیه خارجی این سلولها از روی هم قرار گرفتن تعداد زیادی دیسک شتکیل شده است که غشای پلاسمایی اطراف آن را میپوشاند. رنگدانههای دریافتکننده نور در غشای این دیسکها قرار دارد. ناحیه داخلی سلولهای نوری از تعداد زیادی میتوکندری، هسته و انتهای سیناپسی تشکیل شده است. انتهای سیناپسی این سلولها با نورونهای قطبی شبکیه سیناپس تشکیل میدهد. سلولهای استوانهای در نور کم و سلولهای مخروطی در نور زیاد تحریک میشوند. به علاوه سلولهای مخروطی نسبت به نورهای مختلف تحریک میشوند و به تشخیص رنگها کمک میکنند.
گیرنده بویایی
گیرندههای بویایی نورونهای دوقطبی تمایزیافتهای هستند که از از یک دندریت تشکیل شدهاند. در انتهای این دندریت تعداد زیادی مژک وجود دارد که در مخاط سقف بینی قرار میگیرد. این گیرندهها با تغییر غلظت مولکولهای بو در مخاط بینی تحریک میشوند. در هر بینی میلیونها گیرنده وجود دارد که آکسون آنها با سلولهای میترال سیناپس تشکیل میدهد. عصب بویایی از کنار هم قرار گرفتن آکسون سلولهای میترال تشکیل میشود که پیام عصبی را به پیاز بویایی منتقل میکند. طول عمر گیرندههای بویایی بین ۳۰ روز تا یک سال است و در تنها سلولهای عصبی هستند که به طور مرتب جایگزین میشوند. سلولهای پایه، سلولهای بنیادی تمایزنیافتهای هستند که جایگزین سلولهای بویایی میشوند و سلولهای پشتیبان به حفظ تعادل محیط خارج سلولی نورونها کمک میکنند.
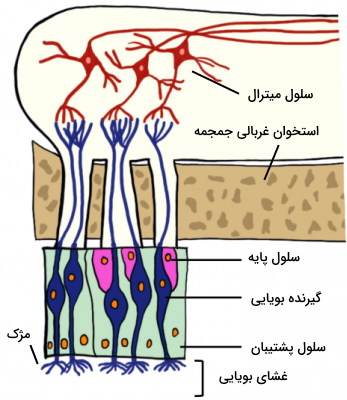
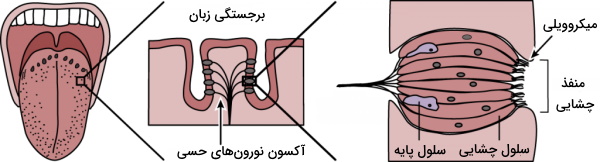
گیرنده چشایی
گیرندههای چشایی، سلولهای پوششی تمایزیافته هستند که در سطح آنها میکروویلی قرار دارد. میکروویلیها مساحت سطح در تماس با مواد غذایی را افزایش میدهند. برجستگیهای فراوان سطح زبان، محل قرار گرفتن جوانههای چشایی است. در هر جوانه چشایی مثل غشای بویایی علاوه بر گیرندهها، سلولهای پشتیبان و پایه نیز وجود دارد. اما این گیرندهها برخلاف گیرندههای بویایی نورون حسی نیستند، بلکه با نورونهای حسی سیناپس میدهند. این گیرندهها به درک مزه شور، شیرین، تلخ، ترش و اومامی (مزه پروتئین) کمک میکنند.
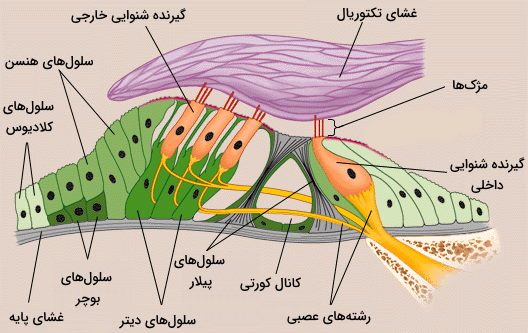
گیرنده شنوایی
گیرندههای شنوایی و تعادل، گیرندههای مکانیکی هستند که در دو بخش متفاوت گوش داخلی (گیرندههای شنوایی در مجرای حلزونی و گیرندههای تعادل در مجاری نیمدایرهای) قرار دارند. این سلولها مثل گیرندههای چشایی، سلولهای اپتلیال تمایزیافتهای هستند که با نورونهای حسی سیناپس تشکیل میدهند. مژکهای سطح این سلول با حرکت مایع اطراف حرکت و باز شدن دریچه کانالها را تحریک میکنند. ساختار گیرندههای تعادل شبیه گیرندههای شنوایی است. اما با تغییر جهت سر تحریک میشوند.
انواع نورون های حرکتی
اگر تا این بخش از مطلب مجله فرادرس با ما همراهی کرده باشید، متوجه شدید سلول عصبی چیست و نورون حرکتی یکی از انواع آن است. نورونهای حرکتی به دو دسته کلی نورونهای حرکتی بالایی و پایینی تقسیم میشوند. نورونهای حرکتی بالایی پیام عصبی قشر مغز را به ساقه مغز یا نخاع منتقل میکنند. نورونهای حرکتی پایینی پیام عصبی نخاع را به ماهیچهها و غدد منتقل میکنند. نورونهای حرکتی حرکات ارادی و غیرارادی بدن را کنترل میکنند. نورون های حرکتی بالایی بیشتر در کنترل حرکات ارادی شرکت میکنند. مسیر هرمی و خارجی هرمی، دو مسیر اصلی انتقال پیام نورونهای حرکتی بالایی به نورونهای حرکتی پایینی است.
در مسیر هرمی، نورونهای حرکتی بالایی از بخشی در بصل النخاع به نام ناحیه هرمی عبور میکنند. نورونهای این مسیر پیام عصبی را از بخش حرکتی در قشر مخ لوب جلویی مغز را به نخاع منتقل میکنند و در کنترل حرکات ارادی نقش دارند. نورونهای حرکتی بالایی در مسیر خارج از هرمی در کنترل تعادل و حرکات غیرارادی بدن نقش دارند. نورونهای حرکتی پایینی به انواع سوماتیک، نورونهای احشایی ویژه و نورونهای احشایی فراگیر (سمپاتیک و پاراسمپاتیک) تقسیم میشوند.
- نورونهای حرکتی سوماتیک: این نورونها از ساقه مغز شروع و به انواع آلفا، بتا و گاما تقسیم میشوند. نورونهای آلفا نقش اصلی را در انقباض ماهیچههای اسکلتی بر عهده دارند و جسم سلولی آنها در ساقه مغز یا نخاع (شاخ جلویی) است. نورونهای حرکتی بتا به انقباض ماهیچههای اسکلتی کمک میکند. نورونهای گاما پیام عصبی را به دوک ماهیچه اسکلتی منتقل میکنند. این نورونها انقباض ماهیچههای اسکلتی را تنظیم میکنند.
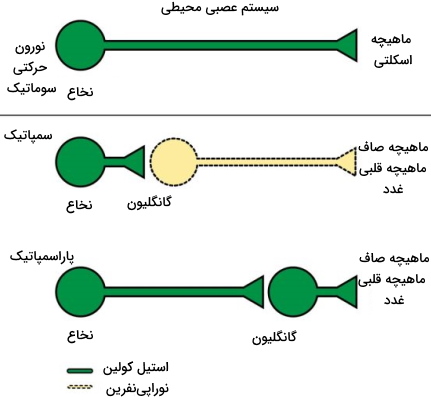
- نورونهای احشایی ویژه: این نورونهای حرکتی در ساقه مغز قرار دارند و پیام عصبی مغز را به ماهیچههای اسکلتی صورت و گردن منتقل میکنند. این نورونها همراه نورونهای حسی هستههای اعصاب جمجمهای V، VII، IX، X و XI را تشکیل میدهند. هستهها محل تجمع جسم سلولی نورونهای سیستم عصبی مرکزی است.
- نورونهای سمپاتیک و پاراسمپاتیک: این نورونها فعالیتهای غیر ارادی بدن را کنترل میکنند و در انقباض ماهیچههای قلب، ماهیچههای صاف دیواره سرخرگها، ماهیچههای صاف دیواره لوله گوارش و ترشح بسیاری از غدد اگزوکرین بدن نقش دارند. نورونهای سمپاتیک و پاراسمپاتیک به انواع پیش گانگلیونی و پس گانگلیونی تقسیم میشوند. گانگلیون محل تجمع جسم سلولی این نورونها است. نورونهای پیشگانگلیونی پیام عصبی را از دستگاه عصبی خارج میکنند و نورونهای پسگانگلیونی این پیام را به اندامها منتقل میکنند. نورونهای سمپاتیک در واکنشهای جنگ و گریز، و نورونهای پاراسمپاتیک در زمان استراحت و گوارش فعال میشوند. تحریک سمپاتیک با افزایش قطر مردمک، کاهش ترشح بزاق، افزایش قطر مسیرهای تنفسی، افزایش ضربان قلب، کاهش قطر رگها، کاهش حرکات دودی لوله گوارش، کاهش خروج آب و سدیم از کلیه، و افزایش ترشح هورمونهای اپینفرین و نوراپینفرین از غدد فوق کلیه همراه است. تحریک پاراسمپاتیک عکس اثر نورونهای سمپاتیک است.
سلول های پشتیبان
در بخشهای قبلی این مطلب از مجله فرادرس توضیح دادیم ساختار و انواع سلول عصبی چیست . اما بافت عصبی از سلولهای دیگری هم تشکیل شده است. سلولهای پشتیبان یا گلیا سلولهای غیرعصبی هستند که به عملکرد نورونها کمک میکنند. آستروسیتها، اولیگودندروسیتها و سلولهای اپندیمال از به عملکرد نورونهای سیستم عصبی مرکزی، و سلولهای شوآن، سلولهای ماهوارهای و سلولهای گلیای روده به عملکرد نورونهای سیستم عصبی محیطی کمک میکنند.
- آستروسیت: آستروسیتها فراوانترین سلولهای پشتیبان سیستم عصبی مرکزی و شبیه ستاره هستند. در مغز و نخاع سلولهای عصبی با مویرگها ارتباط مستقیم ندارند. زاوئد سیتوپلاسی آستروسیتها بین نورونها و مویرگهای خونی ارتباط برقرار میکند. این سلولها غلظت پتاسیم و انتقالدهندههای عصبی در مایع میانبافتی نورونها را تنظیم میکنند. این سلولها بخشی از سد خونی-مغزی هستند.
- اولیگودندروسیت: اولیگودندروسیتها سلولهای میلینساز در سیستم عصبی مرکزی هستند. زوائد سیتوپلاسمی این سلولها دور آکسون نورونهای میپیچد و پوششی لیپیدی به نام غلاف میلین ایجاد میکند. این غلاف سرعت انتقال پیام در آکسون را افزایش میدهد. هر سلول آستروسیت میتواند در تشکیل غلاف پیوندی چند نورون شرکت کند.
- سلولهای اپندیمال: اپندیموسیتها سلولهای مژکدارداری هستند که از کنار هم قرار گرفتن آنها غشای اپندیمال در بطنهای مغز و کانال مرکزی نخاع تشکیل میشود. وظیفه این سلولها ترشح مایع مغزی-نخاعی است. مایع مغزی نخاعی ترکیبی شبیه پلاسمای خون است که بین استخوان جمجمه و بافت عصبی مغز، در بطنهای مغز و بین استخوانهای کانال مرکزی و سلولهای عصبی نخاع قرار دارد. این مایع از مغز و نخاع در برابر فشار فیزیکی محافظت میکند.
- سلولهای شوآن: سلولهای شوآن نقش اولیگودندروسیتها را در سیستم عصبی محیطی بر عهده دارند. برخلاف اولیگودندروسیتها، تمام غشای یک سلول شوآن دور آکسون نورونها میپیچد. در نتیجه بخشهای داخلی غلاف میلین نورونهای در سیستم عصبی مرکزی از لایههای لیپیدی و لایه خارجی آن از هسته و سیتوپلاسم سلولهای شوآن تشکیل شده است. هر سلول شوآن در تشکیل غلاف پیوندی یک آکسون شرکت میکند. به علاوه سلولهای شوآن میکروبها و بقایای سلولی در سیستم عصبی محیطی را فاگوسیتوز میکنند.
- سلولهای ماهوارهای: سلولهای ماهوارهای در گانگلیونهای نورونهای حسی، پاراسمپاتیک و سمپاتیک قرار دارند. این سلولها مثل آستروسیتها غلظت مواد شیمیایی محیط نورونها را تنظیم میکنند.
- سلولهای گلیای روده: این سلولها بین نورونهای سیستم عصبی معده و روده قرار دارند و نقش اصلی آنها حفظ تعادل غلظت مواد شیمیایی در مایع میانبافتی نورونها است.
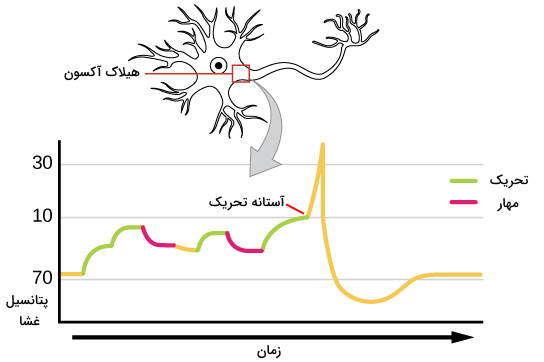
تحریک پذیری سلول عصبی چیست؟
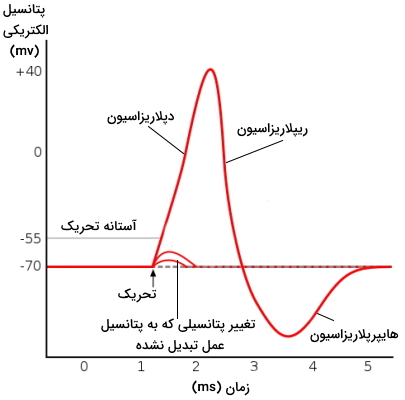
در غشای پلاسمایی نورونها کانالهای یونی زیادی وجود دارد. این کانالها با تغییر نفوذپذیری غشا به یونهای سدیم و پتاسیم در نورون جریان الکتریکی ایجاد میکنند. به این فرایند تحریکپذیری سلول عصبی گفته میشود. قبل از بررسی مراحل تحریکپذیری غشا، بهتر است توضیح دهیم پتانسیل غشای سلول عصبی چیست . فرض کنید یک الکترود در غشای پلاسمایی سلول زنده و الکترود دیگری در مایع میانبافتی این سلول قرار دادهاید و به کمک این الکترودها اختلاف بار یونها یا اختلاف پتانسیل الکتریکی را اندازه میگیرید. عدد بهدست آمده، اختلاف پتانسیل غشای سلول زنده را نشان میدهد و همیشه نسبت به محیط خارج از سلول بیان میشود.
اختلاف پتانسیل غشای نورون در حالت استراحت (وقتی تحریکی وجود ندارد) بین ۳۰- تا ۷۰- میلیولت است. در این حالت بار منفی سیتوپلاسم از مایع خارج سلولی بیشتر و غشا قطبی یا پلاریزه است. اگر پتانسیل غشا مثبتتر (بیشتر از ۳۰-) شود، غشا دپلاریزه و اگر پتانسیل غشا منفیتر شود (۷۰-)، غشا هایپرپلاریزه است.
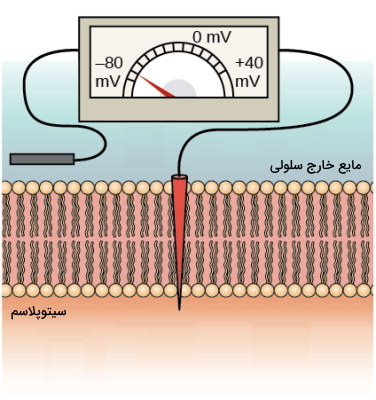
پتانسیل استراحت غشای سلول عصبی چیست؟
پتانسیل استراحت غشا به دلیل اختلاف غلظت یونهای سدیم، پتاسیم و کلر در دو طرف غشا و تفاوت نفوذپذیری غشا به این یونها ایجاد میشود. غلظت پتاسیم و آنیونهای آلی (آمینواسید و پروتئینها) در سیتوپلاسم نورونها و غلظت یون سدیم و کلر خارج از سلول بیشتر است. در این حالت مجموعه غلظت یونهای کاتیون و آنیون در تعادل است. لیپیدهای آبگریز غشا نسبت به یونها نفوذناپذیر است. به همین دلیل انتقال یونها در غشای پلاسمایی نورون و سایر سلولها بهوسیله کانالهای پروتئینی غشا انجام میشود.
بعضی از این کانالها در حالت استراحت نورون باز هستند و به آنها کانالهای همیشه باز یا بدون دریچه گفته میشود. بعضی از این کانالها دریچهدار و در حالت استراحت بسته هستند. این کانالها فقط در پاسخ به پیامهای خارج یا داخل سلولی باز میشوند. یون پتاسیم عامل اصلی تعیین پتانسیل غشا در حالت استراحت است. فرض کنید فقط پتاسیم از غشای نورون عبور میکند و غلظت پتاسیم در سیتوپلاسم بیشتر از محیط اطراف نورون است. اگر کانالهای پتاسیم باز باشد، پتاسیم در جهت شیب غلظت (غلظت کمتر به بیشتر) از سیتوپلاسم خارج میشود.
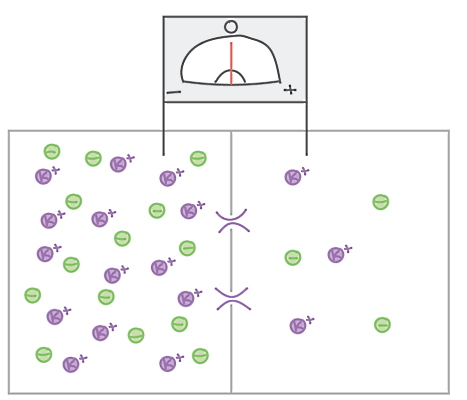
با هر بار خروج پتاسیم بار مثبت خارج از سلول و بار منفی سیتوپلاسم کمی افزایش مییابد. در نتیجه با خروج پتاسیم سیتوپلاسم نسبت به فضای خارج از سلول منفیتر و اختلاف پتانسیل ین دو محیط ایجاد میشود. در این شرایط یونهای منفی سیتوپلاسم با پتاسیم برهمکنش میدهند و از خروج آن جلوگیری میکنند. زمانی که اختلاف غلظت شیمیایی و بار الکتریکی پتاسیم در دو طرف غشا برابر شد، تعداد یونهای پتاسیم خروجی و ورودی به سیتوپلاسم برابر میشود. اگر سلول فقط به یون پتاسیم نفوذپذیر باشد، پتانسیل استراحت غشای نورون برابر با پتانسیل الکتریکی پتاسیم در حالت تعادل است.
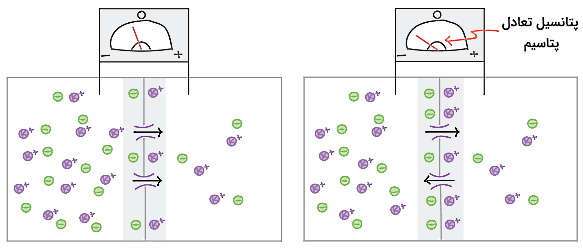
پتانسیل غشای سلولهای پشتیبان با پتانسیل تعادل یون پتاسیم برابر است. اما پتانسیل غشای نورون ها به دلیل نفوذپذیری غشا به یون سدیم از پتانسیل تعادل پتاسیم منفیتر است. فرض کنید غشای نورونها فقط به سدیم نفوذپذیر است. غلظت سدیم خارج از نورون بیشتر از سیتوپلاسم است. به همین دلیل باز شدن کانالها با ورود یون سدیم و مثبتتر شدن سیتوپلاسم همراه است. یونهای منفی با سدیم برهمکنش داده و از ورود آن به سیتوپلاسم جلوگیری میکنند. زمانی که اختلاف غلظت شیمیایی و بار الکتریکی سدیم در دو طرف غشا برابر شد، تعداد یونهای سدیم ورودی و خروجی سیتوپلاسم برابر و پتانسیل تعادل یون سدیم ایجاد میشود.
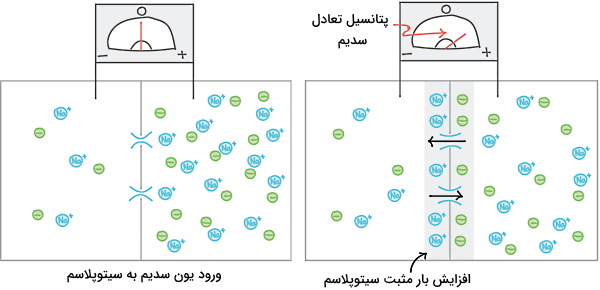
غشای نورونها در حالت استراحت به هر دو یون سدیم و پتاسیم نفوذپذیر است. به همین دلیل پتانسیل استراحت غشای نورون بین پتانسیل تعادل پتاسیم و سدیم است. پمپ سدیم-پتاسیم پتانسیل استراحت را تثبیت میکند. این این پمپ با مصرف هر مولکول ATP سه یون سدیم را به مایع خارج از نورون و دو یون پتاسیم را به سیتوپلاسم منتقل میکند. در نتیجه همیشه غلظت سدیم خارج سلول و پتاسیم داخل سلول بیشتر است.
پتانسیل عمل سلول عصبی
پتانسیل عمل، تغییر موقتی اختلاف پتانسیل بین مایع خارج سلولی و سیتوپلاسم نورون است که به دلیل ورود و خروج ناگهانی یونها ایجاد میشود. برخلاف پتانسیل استراحت، برای ایجاد پتانسیل عمل کانالهای دریچهدار سدیمی و پتاسیمی باز میشود. تغییر اختلاف پتانسیل با باز شدن دریچه این کانالها همراه است. به همین دلیل کانالهای ولتاژی نام دارند. کانالهای ولتاژی سدیمی دو و کانالهای ولتاژی پتاسیم یک دریچه دارند.
انتقالدهندههای عصبی مولکولهای کوچکی هستند که از پایانه آکسون یک نورون ترشح شده و به گیرندههای دندریت نورون دیگر متصل میشوند. اتصال این مولکولها به گیرنده بهطور مستقیم یا غیر مستقیم نفوذپذیری غشا به یونها را تغییر میدهد. اگر پتانسیل غشا به پتانسیل تغییر شکل دریچه فعالکننده کانالهای ولتاژی سدیم (حدود ۵۵- میلیولت) برسد، این کانالها باز و جریانی از یون سدیم وارد سیتوپلاسم میشود. افزایش یون سدیم در سلول با مثبتتر شدن پتانسیل الکتریکی سیتوپلاسم و دپلاریزه شدن غشا همراه است. زمانی که این پتانسیل به پتانسیل تغییر شکل دریچه کانالهای پتاسیمی (حدود ۳۰+ میلیولت) برسد، دریچه کانال ولتاژی پتاسیم باز و دریچه غیرفعالکننده کانال سدیم بسته میشود.
خروج یونهای پتاسیم و توقف ورود یونهای سدیم با منفیتر شدن سیتوپلاسم و ریپلاریزه شدن غشا همراه است. ادامه پیدا کردن خروج یونهای پتاسیم پتانسیل الکتریکی سیتوپلاسم را کاهش (کمتر از ۷۰-) میدهد و غشا هایپرپلاریزه میشود. پمپ سدیم پتاسیم پتانسیل غشا را به حالت استراحت برمیگرداند.
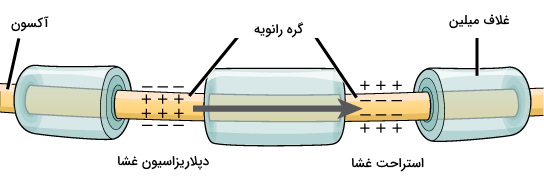
انتقال جریان در سلول عصبی چیست؟
انتقال جریان الکتریکی در نورونها یکطرفه و از دندریت به آکسون است. سرعت انتقال جریان الکتریکی در آکسون به قطر این بخش از نورون و غلاف میلین بستگی دارد. غلاف میلیون آکسون یکپارچه نیست. بین قطعات غلاف میلین بخشی از غشای آکسون با کانالهای ولتاژی سدیمی و پتاسیمی فراوان به نام گره رانویه وجود دارد. پتانسیل عمل فقط در گرههای رانویه ایجاد میشود. در نتیجه سرعت انتقال جریان افزایش مییابد.
سیناپس سلول عصبی چیست؟
سیناپس سلول عصبی محل انتقال پیام عصبی از پایانه آکسون یک نورون به دندریت، جسم سلولی یا آکسون نورون دیگر است. به سلولی که پیام را منتقل میکند، نورون پیشسیناپسی و به سلول عصبی که پیام را دریافت میکند، نورون پسسیناپسی گفته میشود. سیناپسها به دو نوع الکتریکی و شیمیایی تقسیم میشود.
در سیناپسهای الکتریکی غشای دو نورون بهوسیله اتصالات سوراخدار با هم در ارتباط هستند. هر اتصال سوراخدار کانالی غشایی است که از کنار هم قرار گرفتن ۶ پروتئین کانکسین ایجاد شده است. در سیناپسهای الکتریکی پیام عصبی با ورود یونهای سدیم از سلول پیشسیناپسی به سلول پسسیناپسی منتقل میشود. جریان الکتریکی در این سیناپس از سیناپسهای شیمیایی سریعتر منتقل میشود. بیشتر این سیناپسها برخلاف سیناپسهای شیمیایی دوطرفه هستند.
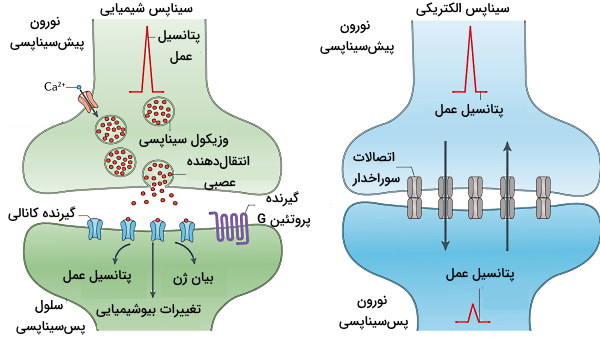
در سیناپسهای شیمیایی غشای سلولی پیشسیناپسی و پسسیناپسی به هم متصل نیست و فاصله بین آنها شکاف سیناپسی نام دارد. در این سیناپسها با رسیدن جریان به انتهای آکسون، کانالهای ولتاژی کلسیم باز و وزیکولهای سیناپسی به غشای پایانه آکسون متصل میشود. در نتیجه مولکولهای کوچکی به نام انتقالدهندههای عصبی از وزیکولها آزاد و به گیرندههای غشایی سلول پسسیناپسی متصل میشود.
آمینواسیدها (گلایسین، گلوتامیکاسید و گاما آمینوبوتیریکاسید)، آمینها (دوپامین، نوراپینفرین، اپینفرین، سروتونین و هیستامین)، نوکلئوتیدها (ATP)، استیل کولین و پپتیدهای کوچک (اندروفین، انکفالین، نوروپپتید P و نوروپپتید K) انواع انتقالدهندههای عصبی هستند. انتقالدهندههای عصبی پپتیدی در هسته جسم سلولی سنتز و به پایانه آکسون منتقل میشود. اما سایر انتقالدهندههای عصبی در انتهای آکسون سنتز میشود.
گیرنده غشای پسسیناپسی کانالهای یونی یا گیرندههای همراه پروتئین G هستند.
- کانالهای یونی: این گیرندهها کانالهای یونی لیگاندی هستند. اتصال انتقالدهنده عصبی به این گیرندهها با تغییر شکل دریچه و ورود یونهای مثبت به سیتوپلاسم همراه است. اگر پتانسیل الکتریکی ایجاد شده از ورود کاتیونها به آستانه تغییر شکل دریچه کانالهای ولتاژی برسد، در سلول پسسیناپسی جریان الکتریکی ایجاد میشود. گیرنده گابا آمینوبوتیریکاسید، گلوتامات، گلایسین در تمام سیناپسها و گیرنده سروتونین، ATP و استیل کولین در بعضی سیناپس از نوع کانال یونی است.
- گیرنده همراه G پروتئین: این گیرندهها پروتئینهایی هستند که ۷ بار از غشا عبور میکنند. جایگاه اتصال به انتقالدهنده عصبی در سطح خارجی و پروتئین G متصل به سطح سیتوپلاسمی است. اتصال انتقالدهنده عصبی به گیرنده با تغییر شکل این پروتئین و فعال شدن پروتئین G همراه است. پروتئین G، آنزیمی است که یکی از فسفاتهای نوکلئوتید گوانین (GTP) را هیدرولیز میکند. پروتئین G فعال آنزیمهای دیگر غشایی را فعال میکند و مولکولهای پیامرسان ثانویه تشکیل میشوند. این مولکولها با ایجاد تغییرات سیتوپلاسمی نفوذپذیری غشا به یونها را تغییر میدهد. ورود کاتیونها سبب مثبت شدن سیتوپلاسم میشود. اگر پتانسیل ایجاد شده بهوسیله این یونها کافی باشد، در سلولی پسسیناسی پتانسیل عمل ایجاد یا مهار میشود. گیرنده دوپامین، نوراپینفرین، هیستامین و نوروپپتیدها در تمام سیناپسها و گیرنده سروتونین، ATP و استیل کولین در بعضی سیناپسها از این نوع گیرنده است.
اتصال انتقالدهنده عصبی به گیرنده ممکن است سبب ایجاد یا مهار پتانسیل عمل در سلول پسسیناپسی شود. بر این اساس سیناپسها را به انواع تحریکی و مهاری تقسیم میکنیم. گلوتامات فراوانترین انتقالدهنده عصبی تحریکی در سیستم عصبی مهرهداران است. گیرنده این انتقالدهنده پیام در بعضی از بخشهای دستگاه عصبی از انواع کانال یونی و در بعضی از قسمتها گیرنده همراه G پروتئین است. اتصال این انتقالدهنده عصبی به گیرنده نفوذپذیری غشا به یونهای سدیم را افزایش میدهد، غشای سلول پسسیناپسی دپلاریزه و پتانسیل تحریکی در سلول ایجاد میشود.
گاما آمینوبوتیریکاسید (GABA) فراوانترین انتقالدهنده عصبی مهاری در سیستم عصبی پستانداران است که در بخشهای مختلف دستگاه عصبی به کانالهای یونی یا گیرندههای همراه G پروتئین متصل میشود. اتصال این انتقالدهنده عصبی به گیرنده با افزایش نفوذپذیری غشا به یونهای کلر همراه است. غلظت کلر خارج از نورونها بیشتر است. به همین دلیل در جهت شیب غلظت وارد سلول میشود و بار منفی سیتوپلاسم را افزایش میدهد. در نتیجه غشا هایپرپلاریزه و ایجاد پتانسیل عمل مهار میشود.
یک نورون همزمان با صدها نورون دیگر سیناپس دارد که ممکن است از انواع مهاری یا تحریکی باشد. پس محرک نهایی برای ایجاد پاسخ سلول عصبی چیست؟ نورون به مجموع تحریکها پاسخ میدهد. دو فرایند تجمیع فضایی و تجمیع زمانی نوع پاسخ نورون را تعیین میکند.
- تجمیع فضایی: در این فرایند مجموع تحریکهایی که همزمان از سیناپسهای مختلف دریافت میشود، ایجاد پتانسیل عمل را مهار یا تحریک میکند. اگر تعداد سیناپسهای مهاری بیشتری باشد، ایجاد پتانسیل عمل مهار و اگر تعداد سیناپسهای تحریکی بیشتر باید، پتانسیل عمل ایجاد میشود.
- تجمیع زمانی: در این فرایند نورون به مجموع پیامهای مهاری و تحریکی ایجاد شده در زمانهای متفاوت پاسخ میدهد. برای مثال هر چه فاصله بین پیامهای تحریکی بیشتر باشد، مجموع تغییر پتانسیل ایجاد شده و احتمال ایجاد پتانسیل عمل بیشتر است.
پایان پیام عصبی
در بخشهای قبلی این مطلب توضیح دادیم که تحریکپذیری سلول عصبی چیست . اما این سوال مطرح میشود که تحریک سلول عصبی چگونه پایان مییابد؟ پایان انتقال پیام عصبی، نورونها را برای دریافت سیگنال بعدی آماده و از خستگی نورون جلوگیری میکند. برای پایانیافتن پیام عصبی در سیناپسهای شیمیایی باید انتقال دهنده عصبی از شکاف سیناپسی خارج شود. یکی از روشهای پایان تحریک عصبی، تجزیه آنزیمی انتقالدهنده عصبی است. برای مثال استیلکولین با این روش از شکاف سیناپسی حذف میشود. آنزیم استیل کولین استراز در غشای سلولهای پسسیناپسی این انتقالدهنده عصبی را به کولین و استات تجزیه میکند. کولین بهوسیله ناقلهای غشایی به نورون پیشسیناپسی برمیگردد و دوباره به استیل کولین تبدیل میشود. در بعضی از سیناپسها انتقالدهنده عصبی بهوسیله ناقل های غشایی به نورون پیشسیناپسی برمیگردد و در بعضی از سیناپسها به سلولهای پشتیبان منتقل میشود.
وظیفه سلول عصبی چیست؟
وظیفه اصلی دستگاه عصبی انتقال پیام، برقراری ارتباط و ایجاد هماهنگی بین دستگاههای مختلف در جهت حفظ تعادل شرایط فیزیولوژیک بدن است. اما سلولهای عصبی در بخشهای مختلف اطلاعات مختلفی را دریافت و پردازش میکنند. برای مثال در بخشهای قبلی توضیح دادیم که نورون های حسی پیام گیرندههای حسی را به بخشهای مختلف مغز منتقل میکنند و نورونهای حرکتی سوماتیک و خودمختار در انتقال پیام عصبی از ساقه مغز و قشر مخ به ماهیچههای و غدد نقش دارند.
جمعبندی سلول عصبی چیست
در این مطلب از مجله فرادرس توضیح دادیم که ویژگی های سلول عصبی چیست. سلولهای عصبی از سلولهای تمایزیافته بدن مهرهداران هستند که بخشهای مختلف دستگاه عصبی مرکزی و محیطی را میسازند. این سلولها از سه بخش دندریت، آکسون و جسم سلولی با شکل و وظیفه متفاوت تشکیل شدهاند. وظیفه دندریتها دریافت پیام عصبی، وظیفه جسم سلولی سنتز پروتئینهای غشایی و آنزیمی، سنتز ATP و انتقال پیام عصبی به آکسون و وظیفه آکسون انتقال پیام به نورون بعدی است. نفوذپذیری غشای نورونها به یونهای پتاسیم، سدیم و کلر منجر به ایجاد پتانسیل عمل یا مهار پتانسیل عمل میشود. انتقالدهندههای عصبی پپتیدهای کوچک، مولکولهای آمینی یا نوکلئوتیدهایی هستند که با اتصال به کانالهای یونی یا پروتئینهای عرض غشایی نفوذپذیری غشا را تغییر میدهند. در بخشهای دیگر مطلب توضیح دادیم که سلولهای عصبی را میتوان بر اساس عملکرد به انواع حرکتی، حسی و بینابینی و بر اساس شکل به انواع دوقطبی، تکقطبی، چندقطبی و دوقطبی کاذب تقسیمبندی کرد.
آزمون سنجش یادگیری
۱. فرایند تمایز سلولی چه نقشی در ایجاد سلول عصبی (نورون) تخصصیافته دارد؟
در تمایز سلولی فقط شکل ظاهری سلولها تغییر میکند، نه عملکرد آنها
تمایز سلولی تنها برای تشکیل سلولهای پوششی کاربرد دارد.
فرایند تمایز، سلولهایی با ساختار و نقش ویژه مانند نورون را پدید میآورد.
تمایز سلولی باعث میشود سلول نقشی ثابت ولی غیرکارکردی داشته باشد.
پاسخ صحیح «فرایند تمایز، سلولهایی با ساختار و نقش ویژه مانند نورون را پدید میآورد» است. در روند تمایز سلولی، هر سلول بسته به وظیفهای که قرار است در بدن ایفا کند، ساختار و نقش ویژهای پیدا میکند. این موجب میشود سلول عصبی (نورون) با ویژگیها و وظیفه تخصصی خود ایجاد شود. گزینههایی چون «تمایز سلولی باعث نقش غیرکارکردی» یا «اختصاصی شدن فقط ظاهر سلول» صحیح نیستند، چون تمایز نه تنها ظاهر بلکه عملکرد سلول را هم تحت تاثیر قرار میدهد. عبارت «تمایز فقط برای سلولهای پوششی» نیز غلط است، زیرا این فرایند انواع مختلف سلولها از جمله نورون را ایجاد میکند.
۲. کدام ویژگی ساختاری نورون بیش از هر چیز آن را از بیشتر سلولهای بدن متمایز میکند؟
وجود سیتوپلاسم و اندامکهای داخلی سلول
وجود غشای سلولی اطراف هسته نورون
داشتن یک هسته مرکزی مثل دیگر سلولها
وجود رشته بلندی به نام آکسون در ساختار نورون
ویژگی «وجود رشته بلندی به نام آکسون در ساختار نورون»، آن را از اغلب سلولهای بدن متمایز میکند؛ چرا که آکسون مسئول انتقال پیام عصبی است و شکل و عملکرد خاصی دارد. «وجود غشای سلولی اطراف هسته نورون»، «وجود سیتوپلاسم و اندامکهای داخلی سلول» و «داشتن یک هسته مرکزی مثل دیگر سلولها»، جزو ویژگیهای اکثر سلولهای بدن بوده و اختصاصی نورون نیستند.
۳. پیام حسی از اندامها به مغز و نخاع توسط کدام ساختار نورون منتقل میشود و این مسیر چه ویژگیای دارد؟
جسم سلولی پیام را بین نورونها رد و بدل میکند و دوطرفه است.
آکسون پیام را از اندامها به مغز منتقل میکند و سریع است.
دندریت پیام را از مغز به ماهیچهها میبرد و یکطرفه است.
شاخههای چندقطبی پیام را از نخاع به اندامها برده و کند عمل میکند.
مسیر انتقال پیام حسی از اندامها به مغز توسط «آکسون» انجام میشود که جریان عصبی را با سرعت زیاد منتقل میکند، زیرا بیشتر آکسونها با غلاف میلین پوشیدهاند. «دندریت» وظیفه دریافت پیام از نورونهای دیگر را دارد، اما پیام را به سوی مغز نمیبرد. «جسم سلولی» نقش پردازش و حمایت را دارد و پیام را بین نورونها رد و بدل نمیکند. «شاخههای چندقطبی» معمولا در نورونهای حرکتی دیده میشوند و پیام را از نخاع به سمت اندام هدف میبرند، نه از اندام به مغز.
۴. در صورتی که غلاف میلین (Myelin sheath) دور آکسون آسیب ببیند، چه اثری بر سرعت انتقال پیام عصبی ایجاد میشود؟
انتقال پیام عصبی بدون تغییر باقی میماند.
سرعت انتقال پیام عصبی کاهش مییابد.
سرعت انتقال پیام عصبی افزایش مییابد.
انتقال پیام عصبی کاملا متوقف میشود.
غلاف میلین نقشی اساسی در افزایش سرعت انتقال پیام عصبی دارد و پیام الکتریکی را به صورت جهشی میان گرههای رانویه سریعتر منتقل میکند. اگر غلاف میلین آسیب ببیند، این هدایت جهشی مختل شده و سرعت حرکت پیام عصبی به طور قابل توجهی کاهش پیدا میکند. عبارت «سرعت انتقال پیام عصبی کاهش مییابد» صحیح است، زیرا وجود میلین کلید انتقال سریع پیامهاست. گزینههایی مانند «سرعت افزایش مییابد» یا «پیام کاملا متوقف میشود» نادرستاند؛ چون نه سرعت بیشتر میشود و نه انتقال به طور کامل متوقف میگردد. همچنین، بیان «انتقال بدون تغییر میماند» اشتباه است، چرا که حذف میلین تاثیر مستقیمی بر کاهش سرعت دارد.
۵. در مسیر عصبی کدام تفاوت اصلی میان نورون حرکتی بالایی و پایینی وجود دارد؟
پایینیها مسئول کنترل هومئوستاز هستند، بالاییها مسئول تعادل حرارتی بدن
پایینیها پیامهای حسی را به مغز میآورند، بالاییها به عضله پیام میفرستند.
بالاییها فرمان حرکت ارادی را منتقل میکنند و پایینیها آن را به عضله میرسانند.
بالاییها فقط در مغز و نخاع هستند، پایینیها فقط بیرون از آنها
نقش اصلی نورونهای حرکتی بالایی در انتقال فرمان حرکت از مغز است و این پیام از طریق مسیر عصبی به نورونهای حرکتی پایینی میرسد. نورونهای حرکتی پایینی پیام نهایی را به عضلات یا غدد مقصد منتقل کرده و باعث اجرای حرکت یا پاسخ فیزیولوژیک میشوند. هیچ شواهدی درباره انتقال پیامهای حسی توسط نورونهای پایینی یا نقش مستقیم این سلولها در کنترل هومئوستاز یا تعادل حرارتی در توضیحات وجود ندارد. همچنین محدود بودن محل قرارگیری بهتنهایی تفاوت عملکردی را توضیح نمیدهد، بلکه اصل تفاوت در نوع ارتباط و نقش انتقال فرمان است.
۶. اگر یکی از انواع کانالهای یونی نورون عملکرد درستی نداشته باشد، چه اثری بر ایجاد پتانسیل عمل خواهد داشت؟
نقش غلاف میلین از بین میرود و پیام کند میشود.
گیرندههای عصبی پیام را به سیناپس ارسال نمیکنند.
جسم سلولی قادر به تولید انرژی لازم برای پیام عصبی نمیشود.
پتانسیل عمل به طور کامل شکل نمیگیرد یا انتقال نمییابد.
در ایجاد پتانسیل عمل، باز و بسته شدن کانالهای یونی (مانند کانال سدیمی و پتاسیمی) برای تغییر سریع پتانسیل غشا ضروری است. اگر هرکدام از این کانالها درست عمل نکند، روند ورود و خروج یونها مختل شده و دیگر پیام عصبی (پتانسیل عمل) شکل نخواهد گرفت یا بسیار ضعیف و ناقص منتقل میشود. گزینههایی مانند «نقش غلاف میلین» یا «جسم سلولی و تولید انرژی» مربوط به بخشهایی دیگر از عملکرد نورون هستند و «گیرندههای عصبی و سیناپس» به ایجاد پتانسیل عمل ارتباط مستقیمی ندارند.
۷. کدام مورد جزو وظایف سلولهای گلیا (glia) در سیستم عصبی مرکزی نیست؟
تامین حمایت ساختاری و حفاظت از نورونها
تنظیم غلظت یونها در اطراف نورونها
تولید غلاف میلین برای تعدادی از اکسونها
انتقال پیام عصبی از طریق پتانسیل عمل (Action Potential)
تنها نورونها قادر به انتقال پیام عصبی از طریق پتانسیل عمل (Action Potential) هستند و سلولهای گلیا این وظیفه را بر عهده ندارند. «تامین حمایت ساختاری و حفاظت از نورونها»، «تنظیم غلظت یونها در اطراف نورونها» و «تولید غلاف میلین برای تعدادی از اکسونها» همگی از وظایف انواع سلولهای گلیا در سیستم عصبی مرکزی محسوب میشوند، اما انتقال پیام عصبی از طریق پتانسیل عمل فقط توسط نورونها انجام میشود.
۸. تفاوت اصلی سیناپس شیمیایی و الکتریکی در چیست؟
امکان تحریک یا مهار نورون بعدی صرفا در سیناپس الکتریکی
وجود اتصال مستقیم سلولی فقط در سیناپس شیمیایی
استفاده از انتقالدهندههای عصبی در سیناپس شیمیایی ولی نه در الکتریکی
انتقال سریعتر پیام عصبی در سیناپس شیمیایی نسبت به الکتریکی
در سیناپس شیمیایی، انتقال پیام با واسطه انتقالدهندههای عصبی (neurotransmitters) صورت میگیرد، اما در سیناپس الکتریکی پیام مستقیما از طریق اتصال کانالی سلول به سلول منتقل میشود و انتقالدهنده شیمیایی دخالت ندارد. بنابراین گزینه «استفاده از انتقالدهندههای عصبی در سیناپس شیمیایی ولی نه در الکتریکی» صحیح است. سرعت انتقال پیام در سیناپس الکتریکی بالاتر است نه شیمیایی؛ اتصال مستقیم سلولی ویژگی سیناپس الکتریکی است نه شیمیایی؛ تحریک یا مهار نورون میتواند در هر دو نوع رخ دهد و مختص سیناپس الکتریکی نیست.
۹. کدام نوع نورون مسئول انتقال پیام حس لمس از پوست به مغز است؟
نورون حسی پیام لمس را به مغز میرساند.
نورون حرکتی دستور را به عضله منتقل میکند.
نورون بینابینی اطلاعات را در مغز پردازش میکند.
نورون گلیا از نورونها پشتیبانی میکند.
انتقال پیام حس لمس از پوست به مغز توسط نورون حسی انجام میشود زیرا نورون حسی اطلاعات محیطی و داخلی مثل لمس را به مغز و نخاع منتقل میکند. «نورون حرکتی» پیامهای مغز را به عضلهها و غدهها میبرد و در انتقال حس نقشی ندارد. «نورون بینابینی» پردازش اطلاعات را میان نورونهای دیگر انجام میدهد و حس لمس را مستقیما منتقل نمیکند. «نورون گلیا» کارش پشتیبانی و تغذیه نورونهاست و ارتباط مستقیمی با انتقال پیام حسی ندارد.
۱۰. گیرندههای نوری (photoreceptors) در چشم چه وظیفهای دارند و چگونه پیام دیداری را آغاز میکنند؟
تغییرات فشار را دریافت و به مغز ارسال میکنند.
مواد شیمیایی را احساس و پیام حسی میفرستند.
امواج صوتی را شناسایی و به پیام الکتریکی تبدیل میکنند.
نور را جذب و آن را به پیام عصبی تبدیل مینمایند.
گیرندههای نوری یا photoreceptors در چشم قادر به جذب نور هستند و با تبدیل این محرک به پیام عصبی، آغازگر مسیر بینایی میشوند. «نور را جذب و آن را به پیام عصبی تبدیل مینمایند» تنها گزینهای است که نقش اختصاصی گیرنده نوری را توصیف میکند. سایر گزینهها به گیرندههای صوتی، فشار یا شیمیایی مرتبط بوده و ارتباطی با عملکرد گیرنده نوری در چشم ندارند.
۱۱. در وضعیت استراحت نورون، به کدام یون غشا بیشترین نفوذپذیری را نشان میدهد و نتیجه این ویژگی بر پتانسیل غشای نورون چیست؟
نفوذپذیری تنها به یون کلر و عدم ایجاد قطبیت خاص
نفوذپذیری بیشتر به یون پتاسیم و ایجاد پتانسیل منفی داخل سلول
نفوذپذیری برابر به هر دو یون و نبود اختلاف پتانسیل
نفوذپذیری بیشتر به یون سدیم و ایجاد پتانسیل مثبت داخل سلول
در حالت استراحت، غشای نورون عمدتا به یون پتاسیم (Potassium) نفوذپذیر است و این باعث تجمع یونهای مثبت خارج سلول و بار منفی نسبی داخل آن میشود. این ویژگی دلیل اصلی ایجاد پتانسیل منفی داخل سلول نسبت به بیرون است. «نفوذپذیری بیشتر به یون سدیم و ایجاد پتانسیل مثبت داخل سلول» نادرست است، زیرا سدیم نفوذپذیری پایینتری دارد. «نفوذپذیری برابر» واقعیت ندارد و همیشه اختلاف پتانسیل وجود دارد. «نفوذپذیری تنها به یون کلر» نیز صحیح نیست و کلر نقش اصلی در پتانسیل استراحت ندارد.
۱۲. اگر نورون بینابینی (interneuron) دچار آسیب شود، چه مشکلی در عملکرد عصبی ایجاد میگردد؟
انتقال پیام از آکسون به دندریت مختل میشود.
پیام عصبی بین نورونهای حسی و حرکتی به درستی پردازش نمیشود.
پیام عصبی از گیرنده حسی نمیتواند به مغز منتقل شود.
حرکت ارادی به طور کلی متوقف خواهد شد.
در صورت آسیب به نورون بینابینی (interneuron)، عملکرد اصلی آن که واسطه و پردازشکنندهٔ پیام عصبی میان نورونهای حسی و حرکتی است مختل میشود. نورونهای بینابینی اطلاعات را دریافت و تحلیل میکنند و جهت ارسال پاسخ مناسب به نورون حرکتی انتقال میدهند. حذف نقش واسطهای آنها باعث میشود ارتباط و پردازش صحیح پیام بین ورودی حسی و خروجی حرکتی انجام نشود. مشکل انتقال پیام از آکسون به دندریت یا توقف کامل حرکات ارادی لزوما نتیجه مستقیم آسیب به این نورونها نیست، و انتقال پیام از گیرنده حسی تا مغز وابسته به زنجیره نورونی کامل، نه فقط این بخش، میباشد.
۱۳. پمپ سدیم-پتاسیم (Sodium-Potassium Pump) چه نقشی در پتانسیل غشایی نورون ایفا میکند؟
خروج یون پتاسیم بیشتر از نورون را تنظیم میکند.
به ورود آزاد یون سدیم به داخل سلول کمک میکند.
یونهای سدیم را از سلول خارج و یونهای پتاسیم را وارد میکند.
توزیع گلوکز درون نورون را کنترل میکند.
پمپ سدیم-پتاسیم با خارج کردن یونهای سدیم و وارد کردن یونهای پتاسیم به داخل سلول نقش اصلی در حفظ تعادل شارژ الکتریکی دو سوی غشاء نورون دارد و این عملکرد پایه ایجاد پتانسیل غشایی است. عبارت «یونهای سدیم را از سلول خارج و یونهای پتاسیم را وارد میکند» بیانگر عملکرد واقعی این پمپ است. گزینههایی مانند «خروج یون پتاسیم بیشتر»، «ورود آزاد یون سدیم» یا کنترل «توزیع گلوکز» نقشی در این فرآیند ندارند و از وظایف پمپ سدیم-پتاسیم نیستند.
۱۴. در نورونها، جمعبندی فضایی و زمانی پیامهای عصبی چه تاثیری بر فرآیند ارسال پیام دارد؟
تعیین میکند آیا پیام نهایی ایجاد و ارسال شود یا نه
باعث حذف کامل همه پیامهای مهاری میشود.
تنها سرعت ورود یونها به نورون را کنترل میکند.
همه پیامهای تحریکی را به صورت خودکار منتقل میسازد.
در نورون جمعبندی فضایی و زمانی پیامهای عصبی نقش کلیدی در تصمیمگیری دارد، زیرا این روند با تجمیع تمام پیامهای تحریکی و مهاری، تعیین میکند که آیا آستانه لازم برای ارسال پیام عصبی (پتانسیل عمل) فراهم شده است یا خیر. بر خلاف «حذف شدن کامل همه پیامهای مهاری» و «انتقال خودکار همه پیامهای تحریکی»، تجمیع پیامها نهایی شدن ارسال یا عدم ارسال پیام را بر اساس جمع اثرها مشخص میکند. همچنین این سازوکار صرفا «سرعت ورود یونها» را کنترل نمینماید بلکه تصمیم نهایی را در ارسال پیام دخیل است.
۱۵. در هنگام نیاز بدن به پاسخ سریع، هدایت جهشی جریان عصبی در آکسون چه برتری نسبت به هدایت پیوسته برای سیستم عصبی دارد؟
افزایش سرعت انتقال پیام عصبی در طول آکسون
تقویت قدرت پیام عصبی در مسیر انتقال
کاهش نیاز به غلاف میلین برای انتقال پیام
کاهش میزان مصرف انرژی توسط سلول عصبی
هدایت جهشی جریان عصبی در آکسون، به دلیل وجود غلاف میلین و گرههای رانویه، باعث افزایش چشمگیر سرعت انتقال پیام در مقایسه با هدایت پیوسته میشود. در این روش، پتانسیل عمل سریعتر از یک گره به گره دیگر میپرد و امکان پاسخدهی آنی سیستم عصبی را فراهم میکند. انتخابهایی مانند «کاهش مصرف انرژی» یا «تقویت قدرت پیام» یا «کاهش نیاز به غلاف میلین» در متن به عنوان ویژگی هدایت جهشی بیان نشدهاند و دلیل برتری این سازوکار همان «افزایش سرعت انتقال پیام عصبی» است.









