



غشای پلاسمایی چیست و ساختار اصلی غشا از چیست؟ – به زبان ساده
غشای سلولی که غشای پلاسمایی نیز نامیده میشود، غشای نازکی است که هر سلول زنده را احاطه کرده و سلول را از محیط اطراف آن جدا میکند. در سلولهای باکتریایی و گیاهی، دیواره سلولی به غشای پلاسمایی در سطح بیرونی آن متصل است. غشای پلاسمایی از یک دو لایه لیپیدی تشکیل شده است که نیمه تراوا است. غشای پلاسمایی حمل و نقل مواد ورودی و خروجی از سلول را تنظیم میکند. در این مقاله به بررسی ساختار غشا و انواع پروتئینهای غشایی، عملکرد غشا و انواع نقل و انتقالات غشایی میپردازیم.

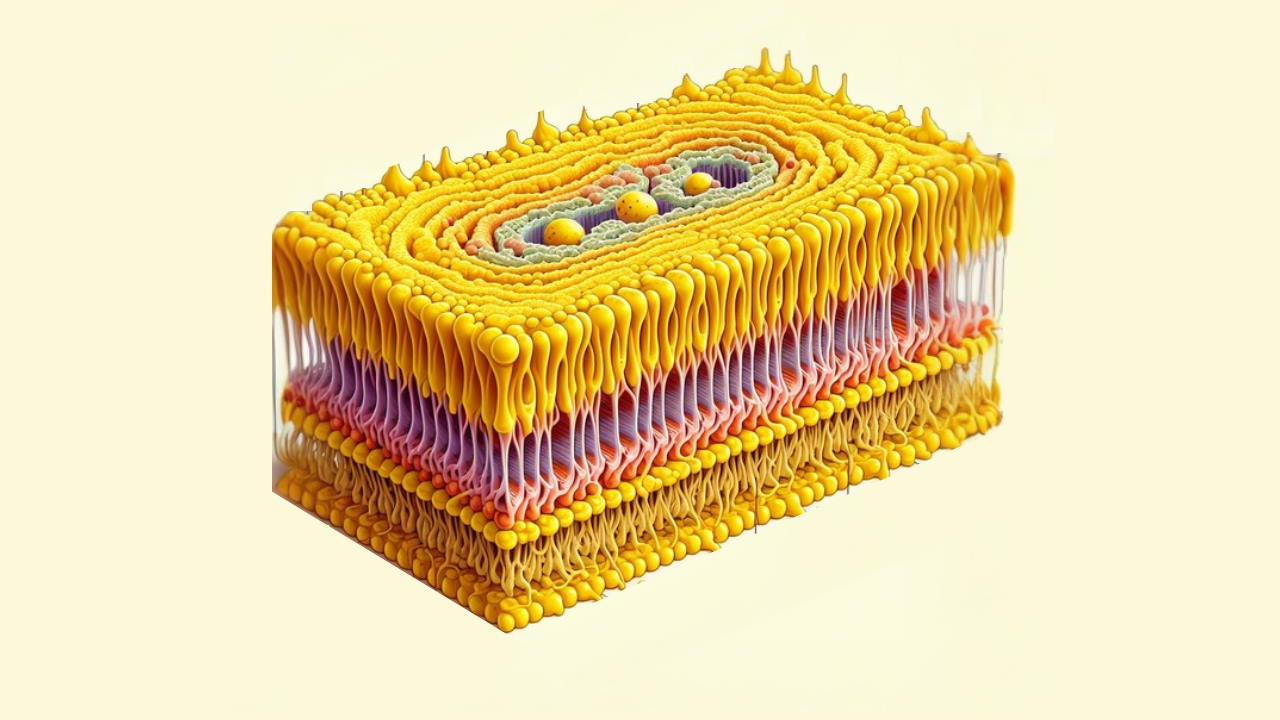


غشای پلاسمایی چیست؟
غشای پلاسمایی سلول و غشاهای پلاسمایی ساختارهایی با ضخامت تقریبی 10 نانومتر هستند که یک مرز محافظ در اطراف سلول و همچنین اندامکهای سلولی تشکیل میدهند. آنها هم مانع از ورود مواد خارجی به سلول میشوند و هم از نشت محتویات سلولی به بیرون جلوگیری میکنند. با ساختاری دولایه لیپیدی که به غشاها خواص فیزیکی و شیمیایی منحصر به فردی میبخشد، این ساختارها همچنین به عملکردهای متنوع و حیاتی سلولی کمک میکنند.
علت نامگذاری غشای پلاسمایی این است که سلول حاوی یک پروتوپلاسم است که ماده زنده نیمه مایع است. این ماده یا پلاسمای زنده در داخل یک غشای بیولوژیکی به نام غشای پلاسمایی وجود دارد. از آنجایی که کل سلول را احاطه کرده است، این غشای پلاسمایی به طور خاص به عنوان غشای سلولی نیز شناخته میشود.

اصطلاح غشای پلاسمایی و غشای سلولی چه تفاوتی دارند؟
همانطور که قبلاً اشاره شد، غشای سلولی یک غشای پلاسمایی است که سلول را احاطه کرده است. اصطلاح غشای پلاسمایی همچنین شامل غشاهای بیولوژیکی میشود که مرز بیرونی بخشهای داخلی (اندامکها) را تشکیل میدهند. بنابراین، به طور دقیقتر، غشای پلاسمایی یک اصطلاح گستردهتر است، زیرا شامل غشاهای دولایه لیپیدی اندامکها، علاوه بر غشای سلولی است.
غشای پلاسمایی و ساختار آن
غشاهای پلاسمایی حاوی انواع مولکولهای بیولوژیکی، به ویژه لیپیدها و پروتئینها هستند. ترکیبات غشا ثابت نیست، اما دائماً برای سیالیت و با توجه به تغییرات در محیط تغییر میکند، حتی در مراحل مختلف رشد سلولی در نوسان است. به عنوان مثال، مقدار کلسترول در غشای سلولی نورون اولیه انسان تغییر میکند و این تغییر در ترکیب بر سیالیت در طول مراحل رشد تأثیر میگذارد. دولایه فسفولیپیدی که یک سد قابل توجه ثابت را در داخل دو مایع (خارج و داخل سلول) تشکیل میدهد یک ساختار اساسی سلول است.
غشای پلاسمایی باید به اندازه کافی انعطافپذیر باشد تا به سلولهای خاصی مانند گلبولهای قرمز و گلبولهای سفید اجازه دهد تا با عبور از مویرگهای باریک تغییر شکل دهند. نسبت پروتئینها، لیپیدها و کربوهیدراتها در غشای پلاسمایی بین انواع مختلف سلولها متفاوت است. با این حال، برای یک سلول انسانی معمولی، پروتئینها حدود 50 درصد از ترکیب جرم را تشکیل میدهند، لیپیدها (از همه انواع آنها) حدود 40 درصد را تشکیل میدهند و 10 درصد باقی مانده از کربوهیدراتها به دست میآیند.

مدل موزائیک سیال
در سال 1972، S. J. Singer و Garth L. Nicolson مدل جدیدی از غشای پلاسمایی را پیشنهاد کردند. در مقایسه با نظریههای قبلی، این نظریه مشاهدات میکروسکوپی و عملکرد غشای پلاسمایی را به بهترین شکل توضیح میدهد. این نظریه مدل موزاییک سیال نامیده میشود. بر طبق مدل موزائیک سیال غشاهای بیولوژیکی را میتوان به عنوان یک مایع دو بعدی در نظر گرفت که مولکولهای لیپیدی و پروتئینی کم و بیش به راحتی در آن پخش میشوند. طبق مدل موزاییک سیال، غشای پلاسمایی موزاییکی از فسفولیپیدها، کلسترول و پروتئینها است که آزادانه و سیال در صفحه غشا حرکت میکنند. به عبارت دیگر، نمودار غشا (مانند تصویر زیر) شامل یک فرآیند پویا است که در آن فسفولیپیدها و پروتئینها به طور مداوم از کنار یکدیگر میلغزند.
سیالیت غشا
ویژگی موزاییکی غشا، که در مدل موزاییک سیال توضیح داده شد، به توضیح ماهیت آن کمک میکند. پروتئینها و سایر اجزای موجود در غشا میتوانند نسبت به یکدیگر حرکت کنند، مانند قایقهایی که روی دریاچه شناور هستند. با این حال، غشا مانند یک بالن نیست که بتواند منبسط و منقبض شود. در عوض، نسبتاً سفت و سخت است و در صورت نفوذ یا اگر یک سلول آب بیش از حد جذب کند، ممکن است بترکد. با این حال، به دلیل ماهیت موزاییکی خود، یک سوزن بسیار ظریف میتواند به راحتی به غشای پلاسمایی نفوذ کند بدون اینکه باعث ترکیدن آن شود، و غشا در هنگام بیرون کشیدن سوزن جریان یافته و خودبخود بسته شود.

ویژگیهای موزاییکی غشا فقط بخشی از سیالیت آن را توضیح میدهد. دو عامل دیگر وجود دارد که به حفظ این ویژگی مایع کمک میکند. یکی از عوامل ماهیت خود فسفولیپیدها است. ساختار دم اسیدهای چرب در هر فسفولیپید میتواند غشا را متراکمتر و سفتتر کرده یا چگالی و انعطافپذیری کمتری ایجاد کند. سیالیت نسبی غشا به ویژه در یک محیط سرد مهم است زیرا یک محیط سرد تمایل دارد از سیالیت غشاها بکاهد و آنها را بیشتر در معرض پارگی قرار دهد. بسیاری از موجودات (مانند ماهیها) با تغییر نسبت انواع مختلف اسیدهای چرب در غشای خود در پاسخ به کاهش دما، قادر به سازگاری با محیطهای سرد هستند.
حیوانات دارای یک غشای اضافی هستند که به حفظ سیالیت کمک میکند. کلسترول که در کنار فسفولیپیدهای غشا قرار دارد، تمایل دارد اثرات دما را بر غشا کاهش دهد. بنابراین، این لیپید به عنوان یک بافر عمل کرده، از مهار سیالیت در دماهای پایین جلوگیری میکند و همچنین از افزایش بیش از حد سیالیت در دماهای بالا جلوگیری میکند. بنابراین، کلسترول، در هر دو جهت محدوده دمایی، عملکرد مناسب غشا و سیالیت آن را گسترش میدهد.
ساختار اصلی غشای پلاسمایی از چیست؟
اجزای اصلی غشای پلاسمایی، لیپیدها (فسفولیپیدها و کلسترول)، پروتئینها و گروههای کربوهیدراتی هستند که به برخی از لیپیدها و پروتئینها متصل هستند. فسفولیپید یک لیپید ساخته شده از گلیسرول، دو دم اسید چرب و یک گروه سر مرتبط با فسفات است. غشاهای بیولوژیکی معمولاً شامل دو لایه فسفولیپید هستند که دم آنها به سمت هم و سر آبدوست به سمت مایع داخل و خارج سلول قرار گرفته است که آرایشی به نام دولایه فسفولیپیدی را میسازد. کلسترول، چربی دیگری که از چهار حلقه کربن ذوب شده تشکیل شده است، در کنار فسفولیپیدها در داخل غشا یافت میشود.
دولایههای لیپیدی عموماً در برابر یونها و مولکولهای قطبی نفوذناپذیر هستند. آرایش سرهای آبدوست و دمهای آبگریز لایه لیپیدی از ورود املاح قطبی (اسیدهای آمینه، اسیدهای نوکلئیک، کربوهیدراتها، پروتئینها و یونها) و از انتشار در سراسر غشا جلوگیری میکند، اما به طور کلی به انتشار غیرفعال مولکولهای آبگریز اجازه میدهد. این به سلول توانایی کنترل حرکت این مواد را از طریق مجتمعهای پروتئینی گذرنده مانند منافذ، کانالها و ناقلها میدهد.
پروتئینهای غشایی ممکن است تا حدی به غشای پلاسمایی گسترش یابند، به طور کامل از غشا عبور کنند، یا به صورت شل به بخش داخلی یا خارجی آن متصل شوند. گروههای کربوهیدرات فقط در سطح بیرونی غشای پلاسمایی وجود دارند و به پروتئینها یا لیپیدها متصل میشوند و گلیکوپروتئینها یا گلیکولیپیدها را تشکیل میدهند. در ادامه هر کدام از این ساختارها را به طور مفصلتر مورد بررسی قرار میدهیم.

فسفولیپیدها و سایر لیپیدها
غشای سلولی از سه دسته لیپیدهای آمفیپاتیک (دارای خاصیت دوگانه آبدوست و آبگریز) تشکیل شده است: فسفولیپیدها، گلیکولیپیدها و استرولها. مقدار هر کدام از این لیپیدها به نوع سلول بستگی دارد، اما در اکثر موارد فسفولیپیدها فراوانترین هستند و اغلب در بیش از 50 درصد از کل چربیهای غشای پلاسمایی نقش دارند. گلیکولیپیدها فقط مقدار کمی حدود 2 درصد را تشکیل میدهند و استرولها بقیه را تشکیل میدهند. در مطالعات نشان داده که غشای گلبول قرمز خون حدود ۳۰ درصد را لیپیدها تشکیل میدهد. با این حال، برای اکثر سلولهای یوکاریوتی، ترکیب غشای پلاسمایی حدود نیمی از لیپیدها و نیمی از پروتئینها از نظر وزنی است.
فسفولیپید چیست؟
فسفولیپیدها که در دولایه قرار گرفته اند، بافت اصلی غشای پلاسمایی را تشکیل میدهند. آنها برای این نقش مناسب هستند زیرا آمفی پاتیک بوده، به این معنی که آنها دارای مناطق آب دوست و آبگریز هستند. این غشا عمدتاً از مولکولهایی به نام فسفولیپیدها تشکیل شده است که به طور خود به خود در یک لایه دوتایی از سر آبدوست خارجی (هیدروفیل) و دمهای آبگریز داخلی (هیدروفوب) قرار گرفتهاند، چنین فعل و انفعالاتی با آب باعث تشکیل غشای پلاسمایی میشود.

هر مولکول فسفولیپید دارای دو بخش است که هر کدام را بررسی کردهایم.
- سر «آبدوست» (hydrophilic): بخش آبدوست یا هیدروفیل یک فسفولیپید سر آن است که حاوی یک گروه فسفات با بار منفی بوده و یک گروه کوچک اضافی که همچنین ممکن است باردار یا قطبی باشد نیز دارد. سرهای آبدوست فسفولیپیدها در دولایه غشایی به سمت بیرون هستند و با مایع آبی داخل و خارج سلول تماس دارند. از آنجایی که مولکولهای آب قطبی هستند، به آسانی برهمکنشهای الکترواستاتیکی (مبتنی بر بار) با سرهای فسفولیپید ایجاد میکنند.
- دم «آبگریز» (hydrophobic): بخش آبگریز یا «هیدروفوب» یک فسفولیپید از دمهای بلند و غیرقطبی اسید چرب آن تشکیل شده است. دم اسیدهای چرب میتواند به راحتی با مولکولهای غیر قطبی دیگر برهمکنش داشته باشد، اما برهمکنش ضعیفی با آب دارد. به همین دلیل، فسفولیپیدها جایی که باید از آب اطراف محافظت شوند از نظر انرژی اینگونه مطلوبتر هستند که دمهای اسیدهای چرب خود را در قسمت داخلی غشا جمع کنند. دولایه فسفولیپیدی که در اثر این فعل و انفعالات ایجاد میشود، مانع خوبی بین داخل و خارج سلول است، زیرا آب و سایر مواد قطبی یا باردار نمیتوانند به راحتی از هسته آبگریز غشا عبور کنند.
به طور جزئیتر یک مولکول فسفولیپید از یک ستون گلیسرول سه کربنی با دو مولکول اسید چرب متصل به کربنهای 1 و 2 و یک گروه حاوی فسفات متصل به کربن سوم تشکیل شده است. این آرایش به مولکول کلی ناحیهای میدهد که به عنوان سر آن توصیف میشود (گروه حاوی فسفات) که دارای ویژگی قطبی با بار منفی است و ناحیهای به نام دم (اسیدهای چرب) که بار ندارد. سر میتواند پیوندهای هیدروژنی ایجاد کند، اما دم نمیتواند.
در آب یا محلول آبی، فسفولیپیدها تمایل دارند خود را طوری مرتب کنند که دمهای آبگریزشان رو به روی یکدیگر و سرهای آب دوستشان رو به بیرون باشد. اگر فسفولیپیدها دم کوچکی داشته باشند، ممکن است یک میسل (یک کره کوچک و تک لایه) تشکیل دهند، در حالی که اگر دمهای بلندتری داشته باشند، ممکن است یک لیپوزوم (یک قطره توخالی از غشای دولایه) تشکیل دهند.
انواع پروتئین های غشایی
پروتئینها دومین جزء اصلی غشاهای پلاسمایی هستند. پروتئینهایی که داخل غشای پلاسمایی تعبیه شدهاند، میتوانند کل عرض غشا را طی کرده یا فقط در یک طرف غشا قرار گرفته باشند. پروتئینها ممکن است به عنوان آنزیم، اتصالات ساختاری برای الیاف اسکلت سلولی یا به عنوان بخشی از مکانهای شناسایی سلول عمل کنند. پروتئینهایی که کل عرض غشا را طی میکنند میتوانند به عنوان کانال یا پمپ جهت انتقال مواد به داخل یا خارج از سلول عمل کنند. دو دسته اصلی از پروتئینهای غشایی وجود دارد که شامل پروتئینهای اینتگرال و پروتئینهای محیطی هستند.

پروتئین های داخل غشایی
«پروتئینهای غشایی اینتگرال» (Integral membrane proteins)، همانطور که از نامشان پیداست، در غشا ادغام میشوند. آنها حداقل یک ناحیه آبگریز دارند که آنها را به بخش آبگریز دولایه فسفولیپیدی متصل میکند. برخی فقط تا حدی به غشا میچسبند، در حالی که برخی دیگر از یک طرف غشا به طرف دیگر کشیده شده و در هر دو طرف در معرض دید قرار میگیرند. پروتئینهایی که در تمام طول غشا امتداد دارند، «پروتئینهای عرض غشایی» ( transmembrane proteins) نامیده میشوند.

پروتئینهای عرض غشایی ممکن است فقط یک بار از غشا عبور کنند یا ممکن است دارای دوازده بخش مختلف غشایی باشند. یک بخش معمولی پوشاننده غشا متشکل از 20 تا 25 اسیدآمینه آبگریز است که در یک مارپیچ آلفا مرتب شدهاند، اگرچه همه پروتئینهای عرض غشایی با این مدل مطابقت ندارند. برخی از پروتئینهای غشایی اینتگرال کانالی را تشکیل میدهند که به یونها یا سایر مولکولهای کوچک اجازه عبور میدهد.
فیبروز کیستیک یک بیماری وابسته به غشای پلاسمایی سلول بوده و به دلیل نقص در پروتئین اینتگرال غشا (که به عنوان یک کانال عمل میکند) ایجاد میشود. پروتئین CFTR یونها را از یک طرف غشا به سمت دیگر حرکت میدهد. هنگامی که به درستی کار نکند، یونها قادر به عبور از غشا نیستند و این امر باعث میشود که مخاط بسیار غلیظی در ریهها و دستگاه گوارش ایجاد شده و منجر به بیماری شود.

پروتئین های غشایی محیطی
«پروتئینهای غشایی محیطی» (Peripheral membrane proteins) در سطوح بیرونی و داخلی غشاها یافت میشوند که به پروتئینهای اینتگرال یا فسفولیپیدها متصل هستند. برخلاف پروتئینهای غشایی اینتگرال، پروتئینهای غشای محیطی به هسته آبگریز غشا نمیچسبند و تمایل بیشتری به اتصال شل دارند.

عملکرد پروتئین های غشایی
پنج نوع مختلف پروتئینهای غشایی که معمولاً با غشای سلولی مرتبط بوده از لحاظ عملکرد متفاوت هستند که هر کدام از آنها در ادامه توضیح داده شدهاند.
گلیکوپروتئین ها
پروتئینهای مسئول شناسایی یا «گلیکوپروتئینها» (Glycoproteins) پروتئینهایی هستند که مولکولهای قندی به آنها متصل هستند. گلیکوپروتئینها به سلولها کمک میکنند یکدیگر را بشناسند. الگوهای منحصربهفرد مولکولهای قند روی سطح سلول را میتواند با گلیکوپروتئینهای مربوطه در سلول دیگر مرتبط باشند. این فرایند شبیه اتصال کلید به قفل بوده و در صورتی که گلیکوپروتئینها مکمل هم باشند، اغلب به یکدیگر متصل میشوند. این فرآیند اتصال به ارتباط بین سلولها منجر شده و این همان چیزی است که به سیستم ایمنی ما کمک میکند تا مهاجمان خارجی را بشناسد و سپس از بدن ما در برابر آنها دفاع کند. برعکس آن نیز صادق است زیرا ویروسها از گلیکوپروتئینها برای اتصال به سلول میزبان و آلوده کردن آن استفاده میکنند.
پروتئین های گیرنده
«پروتئینهای گیرنده» (Receptor Proteins) گیرندههای سطح سلولی (گیرندههای غشایی یا گیرندههای عرض غشایی) گیرندههایی هستند که در غشای سلولها جاسازی شدهاند. گیرنده نوعی پروتئین شناسایی است که میتواند به طور انتخابی یک مولکول خاص را در خارج از سلول متصل کند و این اتصال باعث ایجاد یک واکنش شیمیایی در داخل سلول میشود. لیگاند مولکول خاصی است که به یک گیرنده متصل میشود و آن را فعال میکند. مولکولهای لیگاند ممکن است هورمونها، انتقالدهندههای عصبی، سیتوکینها، عوامل رشد، مولکولهای چسبندگی سلولی یا مواد مغذی باشند. آنها با گیرنده واکنش میدهند تا تغییراتی را در متابولیسم و فعالیت یک سلول ایجاد کنند.
اتصال به گیرنده های غشایی چگونه است؟
ساختار هر گیرنده طوری است که با یک ماده خاص متصل میشود. اتصال یک ماده خاص به گیرنده آن در غشای پلاسمایی میتواند فرآیندهای درون سلولی را فعال کند، مانند فعال کردن آنزیمهای دخیل در مسیرهای متابولیک. این مسیرهای متابولیک ممکن است برای تامین انرژی سلول، ساختن مواد برای سلول، یا تجزیه ضایعات یا سموم سلولی برای دفع حیاتی باشند. به همین ترتیب، هورمونهای خارج سلولی و انتقالدهندههای عصبی به گیرندههای غشای پلاسمایی متصل میشوند که سیگنالی را به داخل سلول به مولکولهای داخل سلولی منتقل میکنند. برخی از سایتهای شناسایی توسط ویروسها به عنوان نقاط پیوست استفاده میشوند. اگرچه آنها بسیار خاص هستند، پاتوژنهایی مانند ویروسها ممکن است برای سوء استفاده از گیرندهها برای ورود به سلول با تقلید از ماده خاصی که گیرنده قرار است به آن متصل شود، تکامل یابند.

پروتئین های کانالی
«پروتئینهای کانالی» (Channel proteins) امکان حرکت مواد را از یک طرف غشا به طرف دیگر بدون نیاز به انرژی فراهم میکنند. مولکول در حال حرکت در یک طرف پروتئین کانال جذب میشود و بدون استفاده از انرژی، مولکول به داخل سلول آزاد میشود. از کانالها برای انتشار تسهیلشده استفاده میشود که مخصوصاً برای مولکولهای قطبی بزرگ و یونهای باردار که به دلیل ماهیت آبگریز دمهای اسید چرب فسفولیپیدهایی که دو لایه را تشکیل میدهند نمیتوانند آزادانه در غشاهای سلولی پخش شوند.
پروتئین های حامل
«پروتئینهای حامل» (Carrier proteins) از انرژی برای حرکت مولکولها در سراسر غشا استفاده میکنند. ماده شیمیایی مورد انتقال ابتدا باید در محل اتصال پروتئین حامل متصل شود. پس از اتصال، حاملها مواد در حال جابجایی را میگیرند و نگه میدارند. سپس، پروتئین حامل تغییر شکل میدهد به طوری که دهانه پروتئین اکنون به سمت دیگر غشای پلاسمایی باز میشود، سپس مواد در حال انتقال در آن سمت غشا آزاد میشود.
کانال های آب
«آکواپورینها» (Aquaporins) که کانالهای آب نیز نامیده میشوند، یک پروتئین عرض غشایی است که به طور انتخابی مولکولهای آب را به داخل و خارج سلول هدایت میکند، در حالی که از عبور یونها و سایر املاح جلوگیری میکند. مولکولهای آب از طریق منافذ کانال در یک مسیر پشت سر هم حرکت میکنند. وجود آکواپورینها باعث افزایش نفوذپذیری غشا در برابر آب میشود. مولکولهای آب از طریق منافذ کانال به صورت تک فایل عبور میکنند. وجود کانالهای آب باعث افزایش نفوذپذیری غشا به آب میشود.

کربوهیدرات های غشا چه هستند؟
کربوهیدراتها سومین جزء اصلی غشاهای پلاسمایی هستند. آنها همیشه در سطح بیرونی سلولها یافت میشوند یا به پروتئینها (در تشکیل گلیکوپروتئینها) یا به لیپیدها (در تشکیل گلیکولیپیدها) متصل میشوند. این زنجیرههای کربوهیدرات ممکن است از 2 تا 60 واحد مونوساکارید تشکیل شده و ممکن است مستقیم یا منشعب باشند. گلیکوپروتئینها و گلیکولیپیدها محلهای ویژهای را روی سطح سلول تشکیل میدهند که به سلولها اجازه میدهد یکدیگر را بشناسند. این عملکرد تشخیصی برای سلولها بسیار مهم است، زیرا به سیستم ایمنی اجازه میدهد بین سلولهای بدن و سلولها یا بافتهای خارجی تمایز قائل شود.
انواع مشابهی از گلیکوپروتئینها و گلیکولیپیدها بر روی سطوح ویروسها یافت میشوند و ممکن است به طور مکرر تغییر کنند و از شناسایی و حمله سلولهای ایمنی به آنها جلوگیری کنند. کربوهیدراتهایی که گلیکوپروتئینها را تشکیل میدهند، مسئول تعیین گروههای خونی A، B، O انسان هستند. این گلیکوپروتئینها توسط سیستم ایمنی تشخیص داده میشوند که منجر به ناسازگاری در گروههای خونی هنگام اهدای خون ناهماهنگ میشود.
ساختارهای اختصاصی غشای پلاسمایی
همه غشاهای پلاسمایی دارای ویژگی نفوذپذیری انتخابی و حاوی پروتئینهای مختلف هستند. با این حال، برخی از غشاها دارای اجزایی هستند که برای یک هدف خاص اختصاصی شدهاند. در ادامه ساختارهای مهم غشای پلاسمایی را بررسی کردهایم.
- میکروویلیها: برآمدگیهای انگشت مانندی روی سطح برخی از سلولها هستند. این برجستگیها افزایشدهنده سطح برای جذب بیشتر هستند. سلولهایی که روده کوچک را میپوشانند حاوی میکروویلی هستند.
- اتصالات: گروه تخصصی دیگری از پروتئینها هستند که به سایر سلولها متصل میشوند. سه نوع اصلی اتصال وجود دارد: اتصالات محکم، دسموزومها و اتصالات شکافدار.
- «اتصالات محکم» (Tight junctions): پروتئینهایی هستند که سلولهای مجاور را خیلی محکم در کنار هم نگه میدارند تا هیچ چیز نتواند بین آنها نفوذ کند. سلولهایی که مجرای گوارشی و ادراری را پوشاندهاند دارای اتصالات محکم زیادی هستند تا اطمینان حاصل شود که محتویات درون آن اندامهای توخالی به لایههای بیرونی یا حفره بدن نشت نمیکنند.
- «دسموزومها» ( Desmosomes): به این ساختارها گاهی اوقات اتصالات لنگر میگویند. این اتصالات سلولها را توسط الیاف به هم نگه میدارند که امکان حرکت بدون جدا شدن را فراهم میکند. سلولهای حاوی دسموزوم در بافت عضلانی و پوست یافت میشوند.
- «اتصالات شکافدار» (Gap junctions): نواحی باز در غشای پلاسمایی هستند که بین دو سلول مجاور یافت میشوند. پروتئینها دو سلول را به هم متصل میکنند در حالی که به مواد شیمیایی اجازه عبور بین سلولها را میدهند.

کار غشای پلاسمایی چیست؟
وظیفه اصلی غشای پلاسمایی محافظت از سلول در برابر محیط اطراف آن است. غشا به صورت نیمه تراوا بوده و موادی که وارد سلول شده و از آن خارج میشوند را تنظیم میکند. سلولهای همه موجودات زنده دارای غشاهای پلاسمایی هستند. در ادامه وظایف غشای پلاسمایی را به ترتیب بیان میکنیم.
سد فیزیکی سلول
غشای پلاسمایی همه سلولها را احاطه کرده و سیتوپلاسم را که ماده تشکیل دهنده سلول است از مایع خارج سلولی موجود در خارج سلول جدا میکند. این امر تمام اجزای سلول را از محیط بیرون محافظت میکند و اجازه میدهد تا فعالیتهای جداگانهای در داخل و خارج سلول اتفاق بیفتد. غشای پلاسمایی از سلول پشتیبانی ساختاری میکند. اسکلت سلولی که شبکهای از رشتههای پروتئینی در داخل سلول است را به هم متصل میکند و تمام قسمتهای سلول را در جای خود نگه میدارد. ارگانیسمهای خاصی مانند گیاهان و قارچها علاوه بر غشا، دیواره سلولی نیز دارند. دیواره سلولی از مولکولهایی مانند سلولز تشکیل شده است. این دیواره پشتیبانی اضافی برای سلول فراهم میکند، و به همین دلیل است که سلولهای گیاهی برخلاف سلولهای جانوری اگر آب بیش از حد در آنها پخش شود، نمیترکند.
نفوذپذیری انتخابی سلول
غشاهای پلاسمایی به طور انتخابی تراوا (یا نیمه تراوا) هستند، به این معنی که فقط مولکولهای خاصی میتوانند از آنها عبور کنند. آب، اکسیژن و دی اکسید کربن به راحتی میتوانند از طریق غشا عبور کنند. به طور کلی، یونها (مانند سدیم، پتاسیم) و مولکولهای قطبی نمیتوانند از غشا عبور کنند. آنها باید به جای انتشار آزادانه از طریق کانالها یا منافذ خاصی در غشا عبور کنند. به این ترتیب، غشا میتواند سرعت ورود و خروج مولکولهای خاص به سلول را کنترل کند.

اندوسیتوز و اگزوسیتوز مواد در سلول
اندوسیتوز زمانی اتفاق میافتد که سلول محتویات نسبتاً بزرگتری را (نسبت به یونها یا مولکولهای منفرد که از کانالها عبور میکنند) میخورد. از طریق اندوسیتوز، یک سلول میتواند مقادیر زیادی مولکول یا حتی باکتری کامل را از مایع خارج سلولی جذب کند. اگزوسیتوز زمانی است که سلول این مواد را آزاد میکند. شکل غشا به خودی خود تغییر میکند تا به مولکولها اجازه ورود یا خروج از سلول را بدهد. همچنین واکوئلها، حبابهای کوچک غشایی را تشکیل میدهد که میتواند مولکولهای زیادی را به طور همزمان منتقل کرده تا مواد را به مکانهای مختلف در سلول منتقل کند.
به دلیل ماهیت غشای پلاسمایی، انواع مختلف سلولها میتوانند مواد مختلفی را وارد سلول کنند. این امر از طریق فرآیندهایی مانند اندوسیتوز، فاگوسیتوز (بیگانه خواری) و پینوسیتوز حاصل میشود. سلولهایی مانند آمیب، ماده یا ذرات غذا (و همچنین سایر میکروارگانیسمها) را احاطه کرده و آنها را همانطور که در داخل سلول هستند میبلعند. این فرآیندها به سلولها اجازه میدهد تا میکروارگانیسمها یا مواد دیگر را تغذیه یا نابود کنند.
سیگنالینگ سلولی
یکی دیگر از وظایف مهم غشا، تسهیل ارتباط و سیگنالدهی بین سلولها است. غشای پلاسمایی این کار را از طریق استفاده از پروتئینها و کربوهیدراتهای مختلف انجام میدهد. پروتئینهای روی سلول، آن را علامتگذاری میکنند تا سایر سلولها بتوانند سلول را شناسایی کنند. غشای پلاسمایی همچنین دارای گیرندههایی است که به آن اجازه میدهد تا زمانی که مولکولهایی مانند هورمونها به آن گیرندهها متصل میشوند، وظایف خاصی را انجام دهد.

نقش در تقسیم سلول
همانطور که گفته شد غشای پلاسمایی یک ساختار پویا است که همیشه در حرکت است. این ویژگی باعث میشود تا در صورت لزوم، تقسیم سلولی آسان شود تا دو سلول دختر از سلول اصلی تشکیل شود. در این شرایط، غشای پلاسمایی در قسمت مرکزی فشرده شده و از هم جدا میشود و دو سلول جدید را تشکیل میدهد.
انواع نقل و انتقالات غشایی
غشای سلولی به عنوان مانعی بین سلول و محیط خارجی، در تنظیم حرکت یا انتقال مولکولها و یونهای مختلف در داخل و خارج سلول نقش دارد. انتقال یا حرکت این مواد از طریق مکانیسمهای انتشار، انتقال فعال و انتقال وزیکولی انجام میشود.

انتشار
اساساً، انتشار شامل حرکت مواد از ناحیهای با غلظت بالا به ناحیهای با غلظت کم است. به این ترتیب، این حالتِ حرکتی تا حد زیادی به عدم تعادل مواد در داخل و خارج سلول بستگی دارد. با توجه به حرکت مواد، انتشار ممکن است به سه شکل باشد. که در ادامه آنها را بررسی کردهایم.
انتشار غیرفعال یا ساده
همانطور که از نام «انتشار غیر فعال یا ساده» (Passive or simple diffusion) پیدا است، این نوع انتشار ساده بوده و شامل حرکت مولکولها از جایی که بسیار پر تراکم به ناحیه کم غلظت است. در بیشتر موارد، این مولکولها کوچک و غیرقطبی هستند. یک مثال خوب برای این امر انتشار گازها (مثلاً اکسیژن) در داخل و خارج سلول است. برخی از مولکولهای دیگری که از غشای سلولی عبور میکنند، مولکولهای آب که بار کمی دارند و دی اکسید کربن هستند.
انتشار تسهیل شده
برخلاف انتشار ساده، «انتشار تسهیلشده» (Facilitated diffusion) فرآیندی است که در آن باید به مولکولها برای عبور از غشا کمک شود. در حالی که این نوع انتشار به گرادیان غلظت وابسته است، این شیوه انتقال به کانالهایی مانند کانالهای پروتئینی نیز وابستگی دارد (مانند آکواپورینهایی که مولکولهای آب را حمل میکنند). مولکولهایی که به این روش انتقال وابسته هستند (ذراتی مانند سدیم و پتاسیم)، بزرگتر و دارای قطبیت هستند. همچنین، انتشار تسهیلشده ممکن است از طریق پروتئینهای حامل رخ دهد. در این شرایط، مولکولها به پروتئینهای حامل متصل میشوند که باعث میشود شکل آن به گونهای تغییر کند که اجازه میدهد مولکولها به داخل یا خارج از سلول منتقل شوند. در این روش انتقال، پروتئینهای حامل ممکن است فقط به یک نوع مولکول معین (خاص) متصل شوند، کانالهای یونی نیز ممکن است در این دسته قرار گیرند. این کانالها اغلب دریچهای هستند و بنابراین فقط نوع خاصی از یون را مجاز به عبور میکنند.

انتقال فعال
برخلاف انتشار، انتقال فعال نوعی انتقال است که در آن مولکولها باید بر خلاف گرادیان خود حمل شوند. به این ترتیب، به گرادیان غلظت وابسته نیست. با توجه به اینکه حمل و نقل فعال شامل حرکت مولکولها بر خلاف گرادیان غلظت است، برای انجام این کار به انرژی نیز نیاز دارد. دو نوع اصلی حمل و نقل فعال وجود دارد که در ادامه توضیح داده شدهاند.
انتقال فعال اولیه
نمونه خوبی از «انتقال فعال اولیه» (Primary active transport) پمپ سدیم/پتاسیم (Na+/K+ pump) است. در اینجا، انتقالدهنده (پروتئین) در غشا تعبیه شده و تمام عرض غشا را طی میکند. به طور معمول، سمت خارج سلولی این ناقل بسته است در حالی که سمت داخل سلولی باز است. سمت داخل سلولی با ATP (یک منبع انرژی) مرتبط است. در این حالت، انتقالدهنده تمایل زیادی به یونهای سدیم واقع در سلول دارد. این یونها وارد ناقل میشوند و به آن متصل میشوند که به نوبه خود باعث میشود ATP فسفات را به انتقالدهنده منتقل کند. این انرژی مورد نیاز انتقالدهنده را برای بستن قسمت باز داخل سلولی و باز کردن انتهای خارج سلولی فراهم میکند.
در نتیجه، تمایل به یونهای سدیم کاهش مییابد و به آنها اجازه میدهد به محیط خارج سلولی رها شوند. هنگامی که این یونها آزاد میشوند، انتقالدهنده تمایل خود را به پتاسیم افزایش میدهد که باعث میشود یونهای پتاسیم وارد شده و به ناقل متصل شوند. به دنبال آن انتهای خارج سلولی بسته شده در حالی که فسفات در انتهای داخل سلولی آزاد میشود. بنابراین، یک ATP جدید به این محل متصل شده و باعث میشود انتهای داخل سلولی باز شود و پتاسیم را به داخل سلول آزاد کند. این نوع حمل و نقل از مقدار قابل توجهی انرژی استفاده میکند. به همین دلیل این ناقلها کم هستند.

انتقال فعال ثانویه
«انتقال فعال ثانویه» (Secondary active transport) در حالی که حمل و نقل فعال ثانویه نیز به پروتئینهای غشایی متکی است، انرژی مورد استفاده در اینجا از گرادیان الکتروشیمیایی یونها به جای ATP مشتق میشود. یکی از پروتئینهایی که در این روش حمل و نقل نقش دارد، انتقال دهنده همراه سدیم - گلوکز 1 است. در ابتدا سمتی از این پروتئین که در تماس با سیتوپلاسم است بسته میشود در حالی که انتهای خارج سلولی باز است. در این حالت دو محل اتصال یون سدیم با بار منفی در معرض دید قرار میگیرند و دو یون سدیم (آنیون سدیم) را جذب میکنند.
با حرکت یونها در شیب الکتروشیمیایی، انرژی آزاد شده و باعث تغییر شکل ناقل پروتئین میشود. به نوبه خود، این تغییر شکل تمایل آن را برای مولکولهای گلوکز واقع در خارج از سلول افزایش میدهد. گلوکز در این محیط غلظت کمتری دارد. افزایش میل ترکیبی برای گلوکز باعث میشود که مولکولهای گلوکز به ناقل متصل شوند و در نتیجه انتهای خارج سلولی پروتئین با باز شدن انتهای سیتوپلاسم بسته شود. با آزاد شدن یونهای سدیم به داخل سلول، میل ترکیبی پروتئین برای گلوکز کاهش مییابد و اجازه میدهد گلوکز به داخل سلول آزاد شود.

انتقالات وزیکول ها
نوع سوم انتقال به عنوان انتقال وزیکولی شناخته میشود و شامل تشکیل وزیکولهای متصل به غشا است. این شامل فرآیندهایی مانند اندوسیتوز و پینوسیتوز است که در آن یک مولکول در تماس با غشای سلولی درونی میشود. به عنوان مثال، در طول اندوسیتوز (اندوسیتوز با واسطه گیرنده)، ماده (مثلاً لیپوپروتئینهای با چگالی کم) ابتدا با گیرندههای خاص واقع در غشای سلولی تماس پیدا میکنند. این امر باعث تحریک تشکیل وزیکول روی غشا از طریق خم شدن غشا به سمت داخل میشود. در این فرآیند، مولکول در وزیکول محصور شده و به داخل سلول منتقل میشود. فرآیندهای مشابه انتقال وزیکولی برای انتقال مولکولهای مورد نیاز سلول، فاگوسیتوز و همچنین تغذیه توسط ارگانیسمهایی مانند آمیب مهم هستند، این روش معمولاً برای انتقال مولکولهای بزرگتر استفاده میشود.










عالی بود و به. زبان ساده تمام مفاهیم رو آموزش می داد به من خیلی کمک کرد