



ماهیچه اسکلتی چیست؟ – به زبان ساده + مکانیسم
ماهیچه اسکلتی یکی از بافتهای اصلی بدن است که ۳۰ تا ۴۰ درصد وزن بدن را به خود اختصاص میدهد. بیشتر ماهیچههای اسکلتی به استخوانهای بدن متصل میشوند و انقباض آنها در حرکت دادن اندامهای مختلف ازجمله دستها، پاها و استخوانهای دنده در تنفس شرکت میکنند. مهمترین ویژگی این ماهیچهها انقباض ارادی آنها است. در این مطلب ساختار ماهیچه اسکلتی، مراحل انقباض آنها، اکتین و میوزین اسکلت سلولی، بیماریهای این بافت و مکانیسم حرکت آنها همراه استخوان را توضیح میدهیم.
- میآموزید ماهیچه اسکلتی چگونه حرکت بدن را ممکن میسازد.
- یاد میگیرید سازوکار انقباض و رهاسازی ماهیچه اسکلتی را در سطح مولکولی تحلیل کنید.
- با انواع فیبرهای عضلانی و منابع انرژی آنها آشنا میشوید.
- تأثیر واکنشهای عصبی بر دقت و قدرت ماهیچه را خواهید آموخت.
- اختلالات ژنتیکی و دارویی مؤثر بر عملکرد ماهیچه اسکلتی را میآموزید.
- تفاوت ماهیچه اسکلتی با قلبی و صاف و کاربرد آن را یاد میگیرید.

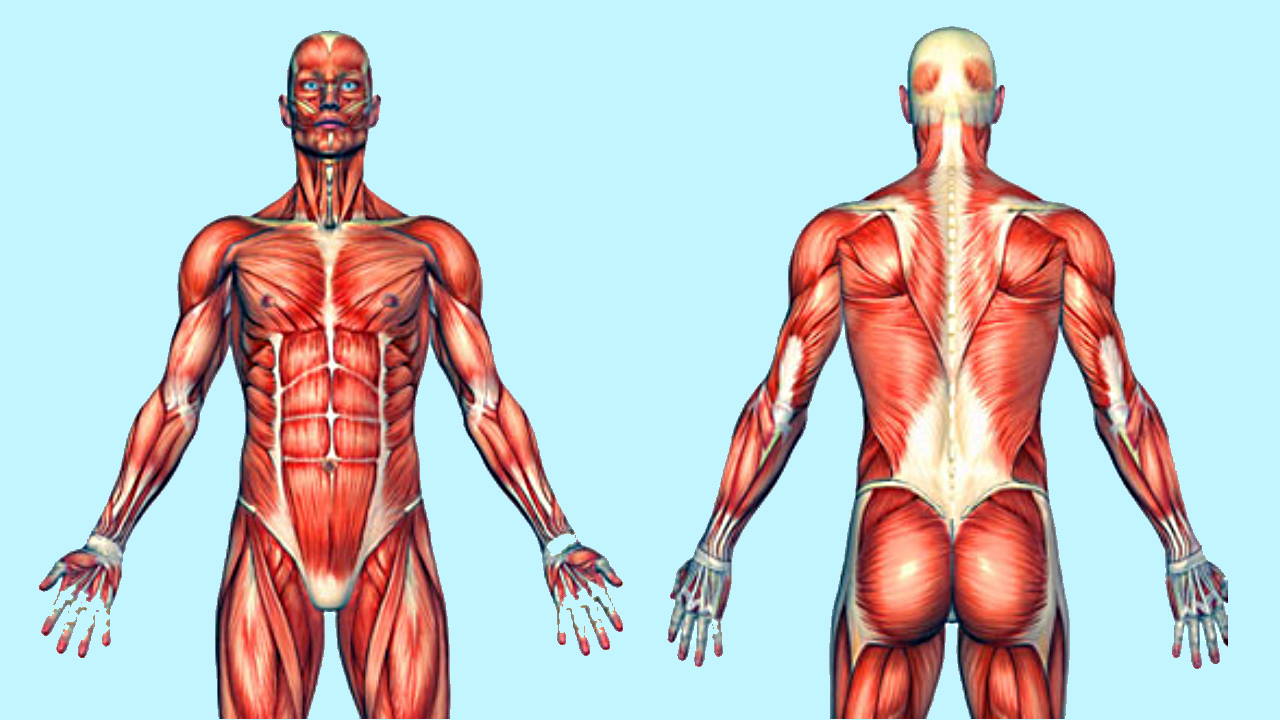


ماهیچه اسکلتی چیست ؟
ماهیچه اسکلتی مجموعهای از سلولهای بسیار تمایزیافته و بخشی از دستگاه حرکتی جانوران است. این بافت به استخوانهای اسکلت داخلی جانوران متصل میشود و انقباض آن به استخوان برای حرکت دادن اندامهای مختلف و حفظ ساختار بدن جانور کمک میکند. به همین دلیل به این بافت ماهیچهای، ماهیچه اسکلتی گفته میشود. این ماهیچه در کنار اسکلت استخوانی به حفظ شکل ایستاده بدن، خم نشدن بر اثر نیروی گرانش و حفظ موقعیت مفصلها کمک میکند. به علاوه ماهیچههای اسکلتی با تولید انرژی گرمایی در پاسخ به گیرندههای دمایی پوست، در تغییر دمای بدن نقش دارند.
تحریکپذیری، انقباض، افزایش یا کاهش طول سلولها و خاصیت کشسانی (الاستیسیته یا انعطافپذیری) چهار ویژگی مهم در بافتهای ماهیچهای ازجمله ماهیچه اسکلتی است.
- تحریکپذیری: نورونها و ماهیچهها سلولهایی هستند که با تغییر پتانسیل الکتروشیمیایی در آنها تحریک میشوند. در این فرایند باز یا بسته شدن کانالهای غشایی سبب تغییر غلظت یونهای مثبت و منفی در سیتوپلاسم سلول و ماترکیس خارج سلولی، در فعالیت سلولها تغییر ایجاد میکند.
- انقباض: فیبرهای پروتئینی درون سلولهای ماهیچهای سبب فشرده و باز شدن این بافت میشود. انقباض این بافتها میتواند تمام اجزای متصل به آنها ازجمله استخوانها را در بدن جابهجا کند.
- تغییر اندازه: شکل سلولهای ماهیچهای شبیه طناب، رشته یا فیبر است. به همین دلیل فشرده یا باز شدن فیبرهای ماهیچهای اندازه این سلولها را تغییر میدهد.
- خاصیت کشسانی: انعطافپذیری ماهیچه اسکلتی سبب میشود سلول پس از کشیده یا فشرده شدن به حالت قبل خود برگردد و تغییر شکل آن دائمی نباشد.
ساختار ماهیچه اسکلتی
هر ماهیچه اسکلتی مجموعهای از فیبرهای ماهیچهای (سلولهای ماهیچه)، نورونهای عصبی، بافت پیوندی و رگهای خونی است. فیبرهای ماهیچهای بخش انقباضی این بافت را تشکیل میدهند که در پاسخ به تحریک عصبی نورونها فعال میشوند. بافت پیوندی از سلولهای ماهیچهای محافظت میکند و فیبرهای الاستیک آن، انعطافپذیری بافت ماهیچه اسکلتی را افزایش میدهد. رگهای خونی این بخش مثل رگهای خونی سایر بخشها، مواد غذایی و اکسیژن لازم برای انجام فرایندهای زیستی را تامین میکند.
بافت پیوندی ماهیچه اسکلتی
تاندون یا زردپی بافت پیوندی بسیار متراکم و محکمی است که بسیاری از ماهیچههای اسکلتی را به استخوان متصل میکند. سه لایه بافت پیوندی (میسیا | Mysia) از تاندون بین سلولهای بافت ماهیچهای گسترش مییابد. این لایههای پیوندی نیروی ایجاد شده از انقباض سلول را به زردپی و استخوان منتقل میکنند. اپیمیسیوم، پریمیسیوم و اندومیسیوم سه لایه بافت پیوندی ماهیجه اسکلتی هستند.
- «اپیمیسیوم» (Epimysium): خارجیترین بافت پیوندی و بخشی است که دور تمام فیبرهای ماهیچهای میپیچید. ساختار این بافت از رشتههای بافت پیوندی نامنظم و متراکم تشکیل میشود. به همین دلیل بسیار محکم و رنگ آن سفید است. اپیمیسیا، بافت ماهیچه اسکلتی را از اندامهای کناری جدا میکند.
- «پریمیسیوم» (Perimysium): بافت پیوندی میانی است که دسته فیبرهای ماهیچهای را از هم جدا میکند. این بافت مثل اپیمیسیوم از رشتههای پیوندی متراکم، محکم و نامنظم تشکیل شده است. فیبرهای ماهیچهای داخل پریمیسیوم با یک رشته عصبی تحریک میشوند. به همین دلیل دستههای ماهیچهای میتوانند جدا از هم منقبض شوند.
- «اندومیسیوم» (Endomysium): داخلیترین بافت پیوندی است که فیبرهای ماهیچهای را از هم جدا میکند. این لایه از رشتههای کلاژنی کمتری تشکیل میشود که یک غشای نازک روی هر سلول یا فیبر ماهیچهای و اطراف ماتریکس خارج سلولی آن تشکیل میدهند. رشتههای این بافت نیروی ایجاد شده از انقباض ماهیچه را به تاندونها منتقل میکنند.

تاندون چیست ؟
تاندون یا زردپی بافت پیوندی است که ماهیچه اسکلتی را به استخوان متصل میکند. تندوبلاستها سلولهای نابالغ تاندون هستند که اندازه و شکل آنها متفاوت است. اما بعد از رشد به سلولهای دوکیشکل و بزرگ تندوسیت تبدیل میشوند. تندوسیتها در سنتز پروتئینهای ماتریکس خارج سلولی و پاسخ به نیرویهای مکانیکی وارد شده به تاندون، نقش دارند. غشای پلاسمایی این سلولها اتصالات شکافدار بسیاری وجود دارد که سرعت انتقال پیام بین دو سلول کنار هم را افزایش میدهد. ماتریکس خارج سلولی این بافت پیوندی از رشتههای کلاژن نوع I و پروتئوگلایکان تشکیل میشود. رشته کلاژن در این بافت ویژگی کشسانی ایجاد میکند و پروتئوگلایکانها ویسکوزیته بافت را افزایش میدهند.
در بعضی قسمتهای بدن، ماهیچه اسکلتی مستقیم به استخوان متصل میشود. در این بافتها اپیمیسیوم بهوسیله بافت پیوندی پریسیستئوم یا پریکندریوم به استخوان متصل میشود.
بافت عصبی ماهیچه اسکلتی
بافت عصبی ماهیچه اسکلتی از نورونهای حرکتی سیستم عصبی سوماتیک تشکیل شده است. هر فیبر ماهیچه اسکلتی با یک نورون حرکتی سیناپس دارد. اما هر نورون حرکتی میتوند با چندین فیبر ماهیچهای سیناپس داشته باشد. به هر نورون حرکتی و تمام فیبرهای ماهیچه اسکلتی که با آن سیناپس دارند یک «واحد حرکتی» (Motor Unit) گفته میشود.

اندازه واحد حرکتی بر اساس تعداد فیبرهای ماهیچهای در بافتهای اسکلتی متفاوت است. برای مثال در ماهیچههای خارج چشمی که کره چشم را حرکت میدهند، هر واحد حرکتی از ۵ تا ۱۰ فیبر ماهیچهای تشکیل شده است که با یک نورون حرکتی سیناپس دارند. در نتیجه کره چشم با انقباض سریع واحدهای حرکتی، حرکت میکنند. واحدهای حرکتی بزرگ در بخشهایی وجود دارند که حرکات ساده بدن مثل حرکت دادن بخشی از بدن خلاف نیروی گرانش را انجام میدهند. ماهیچههای ران و پشت از واحدهای حرکتی تشکیل شدهاند که هر نورون حرکتی با صدها فیبر ماهیچهای سیناپس دارد.
بیشتر بافتهای ماهیچه اسکلتی از ترکیب واحدهای حرکتی کوچک و بزرگ تشکیل شده است. فیبر ماهیچهای واحدهای حرکتی کوچکتر با نورونهای حرکتی در ارتباط هستند که تحریکپذیری بیشتری دارند. تحریک واحدهای حرکتی کوچکتر نیروی کششی کمی در بافت ماهیچهای ایجاد میکند. اگر نیروی بیشتری برای حرکت نیاز باشد، واحدهای حرکتی بزرگ تحریک میشوند. این مدل انقباضی سبب میشود ماهیچه دو سربازو برای بلند کردن یک پَر کمترین نیرو و یک وزنه ۲۰۰ کیلوگرمی نیروی کافی تولید کند و از خستگی مفرط ماهیچه جلوگیری شود.
فیبر ماهیچه اسکلتی
سلولهای ماهیچه اسکلتی یا میوفیبرها، سلولهای استوانهای هستند که طول آنها در بخشهای مختلف بدن تا ۳۰ سانتیمتر و عرض آنها تا ۱۰۰ میکرومتر میرسد. این سلولهای تمایزیافته مثل سایر سلولهای بدن از غشای سیتوپلاسمی، سیتوپلاسم، اندامکها و هسته تشکیل شدهاند. فیبرهای ماهیچه اسکلتی در دوران جنینی از ادغام میوبلاستها تشکیل میشوند. به همین دلیل برخلاف سایر سلولهای بدن چند هسته دارند و انرژی زیادی در پروتئینسازی مصرف میکنند.
در سیتوپلاسم یا سارکوپلاسم این سلولها اندامکهای میتوکندری فراوانی وجود دارد که ATP لازم برای انقباض را فراهم میکند. شبکه سارکوپلاسمی یا اندوپلاسمی این سلولها میزان زیادی یون کلسیم برای انقباض سلول ذخیره میکند و گیرندههای رایودین I در غشای نزدیک به سارکولما در پاسخ به محرکهای محیطی تحریک و از این شبکه به سارکوپلاسم منتقل میشود.
در سارکوپلاسم علاوه بر اندامکها و هسته، تعداد زیادی رشته، تارچه یا میکروفیبریل از جنس پروتئين با قطری حدود ۱٫۲ میکرومتر قرار دارند. این رشتههای پروتئینی با آرایش بسیار منظمی در سارکوپلاسم و در واحدهای مشخصی به نام سارکومر قرار میگیرند. سارکولما غشای پلاسمایی هر فیبر ماهیچهای بخشی است که اندومیسیوم روی آن قرار میگیرد. غشای سارکولما در فواصل مشخص از فیبر ماهیچهای وارد سارکوپلاسم میشود و توبولهای T را تشکیل میدهد. این توبولها نزدیک سیسترنای انتهایی (بخشهای دور از هسته) شبکه سارکوپلاسمی ماهیچه اسکلتی قرار دارند.
پروتئین های ماهیچه اسکلتی
در بخش قبلی توضیح دادیم که انواع رشتههای پروتئینی سارکوپلاسم در واحدهای سارکومری کنار هم قرار میگیرند و واحدهای ساختاری-انقباضی فیبر ماهیچهای را تشکیل میدهند. این رشتهها شامل فیلامنتهای Z، پروتئینهای M، میکروفلامنتهای اکتین، رشتههای میوزین و تایتین میشوند.
- رشتههای Z: رشتههای Z از فیلامنتهای نازک اسکلت سلولی ماهیچه هستند که به سارکولما متصل میشوند. فاصله بین هر دو رشته Z یک سارکومر است. آلفا-اکتینین پرونئین اصلی این رشتهها است.
- تایتین: این رشتههای الاستیک قطر کمی دارند و رشتههای Z یک سارکومر را به رشته Z سارکومر بعدی و میوزینها را به صفحه Z متصل میکنند.
- پروتئین M: این رشتهها از پروتئینهای «میومسین» (Myomesin)، C پروتئین و کراتین کیناز تشکیل شدهاند و در میانه هر سارکومر قرار میگیرند.
- میکروفیلامنتهای اکتین: این رشتههای نازک از یک طرف به صفحه Z متصل هستند و انتهای دیگر آنها نزدیک ناحیه وسط سارکومر قرا میگیرند. برای سنتز این پلیمرها زیرواحدهای پروتئینی G-اکتین (اکتین کروی یا گلوبولی) کنار هم قرار میگیرند و رشتههای F-اکتین (اکتین رشتهای یا فیلامنت) تشکیل میشود.
- میوزین: پروتئین میوزین رشتههای ضخیم پروتئینی هستند که در وسط سارکومر با پروتئینهای M برهمکنش دارند و بهوسیله پروتئینهای تایتین به صفحه Z متصل میشوند. میوزین از دو زنجیره سنگین و چهار زنجیره سبک تشکیل شده است و شکل آن شبیه اسپرم ابه سه بخش سر، گردن و دم تقسیم میشود. دو زنجیره آلفا هلیکسی سبک، ساختار دم و فیلامنتهای ضخیم سارکومر را میسازند. دو زنجیره سبک در بخش گردن فعالیت فسفوریلازی این مولکول را تنظیم میکنند. دو زنجیره سنگین سر، بخشهای اتصالی به میکروفیلامنت اکتین و ATPآز میوزین را تشکیل میدهند. در حالت استراحت ماهیچه اسکلتی سر میوزین به ATP متصل و غیرفعال است.
- نبولین: رشتههای پروتئینی هستند که میکروفلامنتهای اکتین را به صفحه Z متصل میکنند.
- تروپومیوزین: پروتئینهای تروپومیوزین در حالت استراحت ماهیچه اسکلتی، به جایگاه فعال میکروفیلامنتهای اکتین متصل میشوند و از اتصال میوزینها به این رشته جلوگیری میکنند.
- تروپونین: پروتئینی کروی با سه جایگاه فعال است که به میکروفیلامنتهای اکتین (تروپونین I)، تروپومیوزین (تروپونین T) و یون کلسیم (تروپونین C) متصل میشوند.
- دیستروفین: این پروتئین میکروفیلامنتهای اکتین را به پروتئینهای غشایی سارکولما متصل میکند. جهش این ژن منجر به دیستروفی عضله و تغییر شکل آن میشود.

آرایش رشتههای پروتئینی در سارکومر سبب میشود این سلول زیر میکروسکوپ با خطوط تیره و روشن دیده شود. به همین دلیل به این فیبرها ماهیچه مخطط نیز گفته میشود.
- بخش تیره، نوار A یا ناحیه «آنیزوتروپیک» (Anisotropic) بخشی است که فیلامنتهای ضخیم میوزین و فیلامنتهای نازک قرار میگیرند.
- بخش روشن، نوار I یا ناحیه «ایزوتروپیک» (Isotropic) بخشی از فیبر ماهیچهای است که فقط فیلامنتهای نازک اکتین، تروپونین، تروپومیوزین و صفحات Z قرار دارند.
- ناحیه H بخشی در مرکز سارکومر است که فقط فیلامنتهای ضخیم در آن قرا دارند.

مراحل انقباض ماهیچه
ماهیچه اسکلتی در چند مرحله منقبض میشود. در مرحله اول پیام عصبی بهوسیله نورون حرکتی نخاع به این بافت منتقل خواهد شد (مرحله عصبی). در مرحله بعد انتقالدهنده عصبی این نورون به گیرندههای سطح سلول ماهیچه متصل میشوند و اختلاف پتانسیل غشای سلولی را تغییر میدهند (مرحله انتقال پیام عصبی). پس از دپلاریزه شدن سارکولمای ماهیچه اسکلتی و ورود یون کلسیم از شبکه سارکوپلاسمی به سارکولم، فیبرهی پروتئینی حرکت میکنند و ماهیچه منقبض میشود. در نهایت ماهیچه پتانسیل الکتریکی غشا و ماهیچه به حالت استراحت برمیگردد (پایان انقباض و استراحت ماهیچه).
مرحله عصبی انقباض
در بخش قبلی توضیح دادیم که هر فاسیکول ماهیچه اسکلتی (دستههای فیبر ماهیچهای) با یک رشته عصبی در ارتباط است. این رشته عصبی که از شاخه شکمی نخاع خارج میشود و نورون حرکتی سوماتیک نام دارد، در اولین مرحله از انقباض پتانسیل الکتریکی سارکولما را تغییر میدهد. این نورونها بهوسیله پیام عصبی تحریک میشوند که از بخشهای بالایی مغز (در حرکات ارادی قشر مخ) به نخاع میرسد. پتانسیل عمل ایجاد شده در این نورونها بهوسیله باز شدن کانالهای ولتاژی سدیم در طول آکسون انتقال پیدا میکند و به پایانه آکسون میرسد. اختلاف پتانسیل ایجاد شده در پایانه آکسون کانالهای ولتاژی کلسیم را باز میکند. ورود کلسیم به سیتوپلاسم نورون با اتصال این یون به پروتئینهای اگزوسیتوزی (SNARE)، فعال شدن این پروتئینها و اتصال وزیکولهای سیناپسی حاوی انتقالدهنده عصبی (استیل کولین) همراه است.
استیل کولین
استیل کولین انتقالدهنده عصبی اصلی در سیناپس ماهیچههای اسکلتی است. این نروترانسمیتر غیرپروتئینی از کولین و استات در پایانه آکسون نورونهای حرکتی سنتز میشود. کولین یکی از ساختارهای آمینواسید تغییریافته با ساختاری شبیه ویتامین B است که از گوارش مواد غذایی جذب و بهوسیله سیمپورتر سدیم-استیل کولین وارد پایانه آکسون میشود. استات (به شکل استیل کوآنزیم A) یکی از مولکولهای حدواسط چرخه کربس است که در میتوکندریهای فراوان پایانه آکسون حرکتی سنتز میشود.
این دو مولکول در پایانه آکسون بهوسیله آنزیم کولین استیل تراسفراز با هم ترکیب میشوند و با آزاد شدن کوآنزیم A، استیل کولین تشکیل میشود. در غشای وزیکولهای ذخیرهای استیل کولین پمپ پروتونی وجود دارد که با مصرف ATP یون هیدروژن را در خلاف جهت شیب غلظت (غلظت هیدروژن در سیتوپلاسم کمتراست) از سیتوپلاسم پایانه آکسون به فضای داخلی وزیکول سیناپسی منتقل میکند (انتقال فعال اولیه). آنتیپورتر پروتون-استیل کولین برای جبران اختلاف الکتروشیمیایی ایجاد شده بهوسیله پمپ، با انتقال یک پروتون به سیتوپلاسم، یک مولکول کولین را به وزیکول منتقل میکند (انتقال فعال ثانویه).

پروتئین غشایی سیناپتوتاگمین و سیناپتوبروین در غشای وزیکولهای سیناپسی و پروتئینهای SNAP25 و سینتاکسین در غشای پایانه آکسون به خروج استیل کولین از پایانه کمک میکنند. یون کلسیم سیتوپلاسم سبب اتصال پروتئین سیناپتوتاگمین وزیکول به SNAP25 غشا و سیناپتوبروین وزیکول به سینتاکسین غشای پایانه آکسون میشود. اتصال این پروتئینها سبب نزدیک شدن دو غشا به هم و در نهایت ادغام آنها میشود. در نتیجه استیل کولین از پایانه آکسون نورون حرکتی وارد غشای سیناپسی خواهد شد. استیل کولین استراز آنزیمی در غشای پایانه آکسون است که استیل کولین را به کولین و استات تجزیه و تحریک ماهیچه اسکلتی را متوف میکند.
مرحله انتقال پیام عصبی
سارکولمای سلولهای ماهیچه اسکلتی تاخوردگیهایی دارد که مساحت غشا را افزایش میدهد. در نتیجه کانالهای یونی دریچهدار بیشتری در این غشا قرار میگیرند. در ساختار این کانالهای حساس به لیگاند، یک جایگاه اتصالی به استیل کولین وجود دارد. اتصال استیل کولین به کانالهای لیگاندی سبب تغییر کنفورماسیون زیرواحدهای پروتئینی این کانال و باز شدن آن میشود. در نتیجه یون سدیم در جهت شیب غلظت از ماتریکس خارج سلولی وارد سیتوپلاسم و پتاسیم از در جهت شیب غلظت از سیتوپلاسم خارج میشود. اما تعداد سدیمهایی که وارد سیتوپلاسم میشوند از پتاسیمهای خروجی سیتوپلاسم بیشتر است. به همین دلیل پتانسیل الکتریکی سارکولما افزایش مییابد و مثبتتر میشود (پتانسیل صفحه انتهایی | End Plate Potential). اگر پتانسیل صفحه انتهایی به آستانه تحریک (حدود ۵۵- mV) برسد، کانالهای ولتاژی سدیم باز، جریانی از یون سدیم وارد سارکوپلاسم و پتانسیل الکتریکی سارکولما ۳۰+ mV میشود (دپلاریزاسیون غشا).

پتانسیل الکتریکی ایجاد شده در طول فیبر ماهیچه اسکلتی حرکت و در توبولهای T کانالهای کلسیمی حساس به ولتاژ (کانالهای نوع L کلسیم یا گیرنده دیهیدروپیریدین) را فعال میکند. فعال شدن این گیرندهها با اتصال آن به گیرندههای رایودین و خارج شدن رادنودین از غشای سارکوپلاسمی همراه است. در نتیجه جریانی از یون کلسیم وارد سارکوپلاسم و به تروپونین متصل میشود.
حرکت اکتین و میوزین ماهیچه اسکلتی
اتصال کلسیم به تروپونین C مولکول تروپونین و تروپومیوزین متصل به آن را بالا میکشد. در نتیجه جایگاه اتصال اکتین در دسترس سر میوزین قرار میگیرد. همزمان هیدرولیز ATP در سر میوزین سبب فسفوریله شدن زنجیره سنگین، جلو کشیده شدن (سمت خطوط Z) سر مولکول و اتصال به اکتین میشود. در این حالت سر اکتین به فسفات معدنی (Pi) و مولکول ADP متصل است. هیدرولیز فسفات معدنی سبب عقب کشیده شدن سر میوزین و اکتین متصل به آن (سمت خط M) میشود. تبادل ADP با ATP سبب جدا شدن میوزین از اکتین میشوند. اتصال میوزین به اکتین و کشیده شدن این میکروفیلامنت به مرکز سارکومر تا زمانی ادامه دارد که دو رشته اکتین کاملا همپوشانی داشته باشند. در این حالت مولکولهای تایتین پروتئینهای صفحه Z را سمت مرکز سارکومر میکشند. در نتیجه طول ماهیچه کاهش مییابد.

با همپوشانی میکروفیلامنتهای اکتین در انقباض و کشیده شدن صفحات Z بهوسیله پروتئینهای تایتین، اندازه خطوط میکروسکوپی را تغییر میدهد. بنابراین در انقباض ماهیچه اسکلتی ناحیه H ناپدید میشود. اندازه باند B (خط تیره) در هر سارکومز ثابت است اما فاصله بین باندهای B بین دو سارکومر کناری تغییر میکند و اندازه باند I (خط روشن) کاهش مییابد.

پایان انقباض و استراحت ماهیچه اسکلتی
در غشای ساکولما علاوه بر کانالهای ولتاژی سدیم، تعداد بسیار زیادی کانال ولتازی پتاسیم وجود دارد. دریچه فعالکننده و مهارکننده این کانالها به ولتاژ دپلاریزاسیون غشا (۳۰+ میلیولت) حساس هستند. در نتیجه دپلاریزاسیون غشا سرعت باز شدن دریچههای فعالکننده بیشتر سرعت بسته شدن دریچههای مهارکننده است. با باز شدن کانال یون پتاسیم با سرعت و در جهت شیب غلظت به بیرون منتقل میشود. جریان یون پتاسیم به خارج سلول با کاهش بار الکتروشیمیایی مثبت در سارکولم، منفی شدن داخل سلول (ریپلاریزاسیون) و بسته شدن آهسته کانالهای پتاسیمی همراه است. در این حالت پتانسیل الکتریکی غشا به حالت قبل از استراحت (۹۰- میلیولت) میرسد و پمپ سدیم-پتاسیم با انتقال سه یون سدیم از سارکوپلاسم به ماتریکس و ۲ یون پتاسیم از ماتریکس به سارکوپلاسم، پتانسیل الکتریکی غشا در حالت استراحت را ثابت نگه میدارد.
زمانی که پتانسیل غشا به مثبتترین میزان خود (۳۰+ میلیولت) میرسد، کلسیم از تروپونین جدا میشود. در نتیجه تروپومیوزین به جای قبلی خود برمیگردد و جایگاه اتصال به میوزین در اکتین را میپوشاند. پمپ کلسیم-پروتون و آنتیپورتر سدیم-کلسیم یون کلسیم را از سارکوپلاسم به شبکه سارکوپلامی منتقل میکنند.
- پمپ کلسیم-پروتون یکی از پروتئینهای عرض غشایی است که با هیدرولیز ATP یک یون کلسیم را خلاف جهت شیب غلظت وارد شبکه سارکوپلاسمی و پروتون را از این اندامک خارج میکند (انتقال فعال اولیه).
- آنتیپورتر سدیم-کلسیم یکی از ناقلهای پروتئینی غشای شبکه سارکوپلاسمی است که یون سدیم را در جهت شیب غلظت از این اندامک خارج میکند و از انرژی الکتروشیمیای ایجاد شده برای انتقال یون کلسیم در خلاف جهت شیب غلظت به لومن شبکه سارکوپلاسمی استفاده میکند (انتقال فعال ثانویه).
همزمان آنتیپورترهای سدیم-کلسیم و پمپ پروتون-کلسیم در غشای ساکولما یون سدیم و پروتون را در جهت شیب غلظت وارد ساکوپلاسم فیبر ماهیچه اسکلتی و از انرژی الکتروشیمیایی آن و هیدرولیز ATP برای انتقال کلسیم خلاف شیب غلظت (انتقال از سارکوپلاسم به ماتریکس خارج سلولی) استفاده میکنند. در این حالت رشتههای پروتئینی به حالت قبل از انقباض برمیگردند و ناحیه H زیر میکروسکوپ دیده میشود.
سنتز ATP در ماهیچه اسکلتی
در بخش قبلی توضیح دادیم که اتصال سر میوزین به جایگاه فعال اکتین وابسته به مولکول ATP است. به علاوه پمپهای غشای پلاسمایی و شبکه سارکوپلاسمی بردون هیدرولیز ATP کلسیم را بین لومن شبکه سارکوپلاسمی، سارکوپلاسم و ماتریکس خارج سلولی منتقل نمیکنند. به همین دلیل انقباض ماهیچه و بازگشت به حالت استراحت بدون غلظت کافی ATP در سلول ایجاد نمیشود. متابولیسم کراتین فسفات، گلیکولیز، تخمیر و تنفس هوازی چهار مکانیسم اصلی سلولهای ماهیچه اسکلتی برای تامین ATP است.
متابولیسم کراتین فسفات
کراتین یکی از آمینواسیدهای تغییریافته بدن و مولکول ذخیرهای در ماهیچهها و نورونهای مغز است. در زمان استراحت ماهیچه، فسفات معدنی از ATP به کراتین منتقل شده و کراتین فسفات و ADP تولید میکند. کراتین فسفات انرژی شیمیایی را به شکل پیوندهای فسفری ذخیره میکند. با تحریک فیبر ماهیچه اسکلتی و شروع انتقباض، آنزیم کراتین کیناز با انتقال فسفات از کراتین به ADP، مولکول ATP لازم برای اتصال سر میوزین به اکتین را فراهم میکند.
گلیکولیز
کراتین فسفات ATP های لازم برای ۱۵ ثانیه اول انقباض ماهیچه را فراهم میکند. پس از آن سلول با تجزیه گلوکز در فرایند گلیکولیز ATP لازم برای انقباض را تامین میکند. گلیکولیز یک مسیر آنزیمی بیهوازی است که شروع و پیشرفت آن به وجود اکسیژن وابسته نیست. گلوکز ورودی در این فرایند از خون به سارکوپلاسم سلول منتقل یا با شکسته شدن مولکولهای گلیکوژن ذخیرهای در این بافت تامین میشود. از هر مسیر گلیکولیز ۲ مولکول ATP و دو پیروویکاسید تولید میشود. اگر اکسیژن کافی در بافت ماهیچهای وجود داشته باشد، پروویکاسید به میتوکندری منتقل خواهد شد تا در واکنشهای زنجیره انتقال الکترون و تولید ATP شرکت کند. اما در شرایط بیهوازی (غلظت اکسیژن پایین) پیروویکاسید به لاکتیک اسید تبدیل میشود تا ATP در واکنشهای تخمیری تولید شود.
تخمیر
در تمرینات سخت ورزشی که اکسیژن کافی به بافتهای ماهیچه اسکلتی منتقل نمیشود، سلول از مسیر پیرووات-لاکتیکاسید برای تبدیل NADH (مولکول مورد نیاز در گلیکولیز) به استفاده میکند. به این ترتیب مسیر گلیکولیز ادامه مییابد. اما از آنجایی که تنها دو مولکول ATP در این مسیر سنتز میشود، انرژی کافی سلول تامین نخواهد شد.
تنفس هوازی
تنفس هوازی کارآمدترین مسیر تولید ATP در فیبرهای ماهیچه اسکلتی است و حدود ۹۵٪ ATP سلول بهوسیله این فرایند تامین میشود. در تنفس هوازی یا تنفس سلولی پیروویکاسید تشکیل شده از گلیکولیز پس از ورود به میتوکندری و شرکت در زنجیره انتقال الکترون به کربن دیاکسید، آب و ATP تبدیل میشود. از هر مولکول وارد شده در این مسیر ۳۶ مولکول ATP تولید میشود.
انواع فیبر ماهیچه اسکلتی
فیبرهای ماهیچه اسکلتی مختلفی در هر فاسیکولها قرار دارند که ساختار، فعالیت و متابولیسم آنها با هم متفاوت است. در این بخش فیبرهای نوع I، فیبر نوع IIA و فیبر نوع IIX را بررسی میکنیم. تعداد این فیبرهای ماهیچهای در هر فاسیکول بهوسیله ویژگیهای ژنتیکی هر فرد و استرسهای محیطی تعیین میشود.
فیبر ماهیچهای نوع یک
یکی از ویژگیهای مهم فیبرهای ماهیچه نوع I یا فیبر ماهیچهای قرمز سرعت پایین انقباض و تامین انرژی در واکنشهای اکسیداسیون است. قطر فیبر ماهیچهای در این سلولها کمتر از فیبرهای IIA و IIX است. تعداد مویرگهای خونی در این فیبرها بسیار زیاد است. در نتیجه خون با غلظت اکسیژن بالا سبب رنگ فیبرهای ماهیچه اسکلتی نوع I قرمز روشن میشود. به علاوه آهن موجود در پروتئینهای میوگلوبین این فیبر که مثل هموگلوبین خون مولکول اکسیژن را ذخیره میکند، در قرمز دیده شدن فیبرهای نوع I نقش دارد.
تعداد میتوکندریهای این فیبر ماهیچهای مثل مویرگها بسیار زیاد است تا انرژی (ATP) لازم برای انقباض ماهیچه را در فرایند تنفس هوازی فراهم کند. مولکول ذخیره انرژی در این فیبرهای ماهیچهای تریگلیسرید است. از بتا اکسیداسیون این مولکول در میتوکندریها، گلیسرول، اسیدچرب و ATP تولید میشود.
سر میوزین در این فیبرهای ماهیچهای فعالیت ATP هیدرولازی پایینی دارد به همین دلیل سرعت انقباض در این فیبرهای ماهیچهای از فیبرهای IIA وIIX بسیار کمتر است. در نتیجه فیبرهای این ماهیچه نسبت به خستگی انقباض مقاوم هستند و هر انقباض آنها ممکن است تا چند ساعت طول بکشد. اما قدرت انقباضی آنها از فیبرهای IIA وIIX کمتر است. به همین دلیل تعداد فیبرهای نوع I در بافتهای ماهیچهای حفظ شکل و موقعیت بدن بیشتر است. به علاوه پیام عصبی نورونهای حرکتی اول به این فیبرها انتقال مییابد.
فیبر ماهیچه ای نوع IIA
ویژگی اصلی این فیبرهای ماهیچه اسکلتی سرعت زیاد انقباض و تامین انرژی بهوسیله تنفس هوازی است. قطر فیبرها، تعداد میتوکندریها، مویرگهای خونی و مولکولهای ذخیرهای گلیکوژن در این فیبرها بیشتر از فیبرهای I و IIX است. از آنجایی که مویرگهای خونی تمام بخشهای این ماهیچه قطور را تغذیه میکنند و فاصله آنها با هم بیشتر از فیبرهای ماهیچهای نوع I است، رنگ فیبرهای ماهیچهای IIA صورتی دیده میشود. قدرت انقباضی این فیبرها از فیبرهای نوع I بیشتر اما از فیبرهای IIX کمتر است و آنزیم کراتین کیناز موجود در این سلولها با فسفوریلاسیون کراتین به تامین ATP در شروع انقباض کمک میکند. مقاومت این فیبرها نسبت به خستگی مفرط کمتر از فیبر نوع I و معمولا حدود ۳۰ دقیقه است. این فیبرهای ماهیچهای بیشتر در بافتهای اسکلتی قرار دارند که برای راه رفتن و دویدن سریع منقبض میشوند. انجام تمرینات ورزشی استقامتی مثل ماراتن در زمان طولانی، منجر به افزایش مویرگهای خونی، میتوکندریها و میوگلوبین در این فیبر ماهیچهای و تبدیل آن به فیبر ماهیچهای نوع I میشود.

فیبر ماهیچه ای نوع IIX
ویژگی اصلی فیبرهای ماهیچه اسکلتی نوع IIX سرعت زیاد انقباض و تامین انرژی بهوسیله گلیکولیز است. قطر این فیبرهای ماهیچهای از فیبرهای نوع II بیشتر و از IIA کمتر است. تعداد مویرگهای خونی این فیبر ماهیچه اسکلتی از فیبرهای II و IIA بسیار کمتر است. به همین دلیل به آنها فیبر ماهیچهای سفید نیز گفته میشود و رنگ آن صورتی بسیار روشن است. به علاوه تعداد میوگلوبینهای این فیبر از دو نوع دیگر بسیار کمتر است.
تعداد ATP تولید شده در فرایندهای هوازی بسیار بیشتر از فرایندهای بیهوازی است. اما زمان بیشتری برای تجزیه گلوکز و تولید ۳۶ ATP نیاز دارند. فیبرهای نوع IIX ماهیچههایی هستند که در حرکات سریع بدن منقبض میشوند و زمان کمی برای تامین ATP دارند. به همین دلیل تمام ATP مورد نیاز خود را بهوسیله فرایندهای بیهوازی (گلیکولیز و کراتین فسفات) تامین میکنند. در نتیجه تعداد اندامکهای میتوکندری در این سلولها بسیار کمتر از فیبرهای نوع I و IIA اما ذخیره گلیکوژن بسیار بیشتر است. قدرت انقباضی فیبر ماهیچهای نوع IIX از دو فیبر ماهیچه دیگر بیشتر اما مقاومت آن به خستگی مفرط بسیار کمتر (کمتر از ۱ دقیقه) است. انجام تمرینات ورزشی استقامتی مثل وزنهبرداری با افزایش سنتز میوفیبریلها برای افزایش قدرت انقباض ماهیچه اسکلتی و تبدیل فیبرهای ماهیچهای I به IIX همراه است. تفاوت انواع فیبرهای ماهیچه در جدول زیر خلاصه شده است.
| فیبر نوع I | فیبر نوع IIA | فیبر نوع IIX | |
| فعالیت میوزین ATPase | آهسته | سریع | سریع |
| قطر فیبر | کوچک | متوسط | بزرگ |
| زمان انقباض | طولانی | کوتاه | کوتاه |
| فعالیت پمپ سارکوپلاسم | آهسته | سریع | سریع |
| مقاومت به خستگی | زیاد | زیاد | بسیار کم |
| منبع انرژی | هوازی | هوازی و بیهوازی | بیهوازی |
| تعداد میتوکندری | زیاد | متوسط | کم |
| رنگ بافت | قرمز | قرمز | سفید |
| میوگلوبین | دارد | دارد | ندارد |
| تراکم مویرگ | زیاد | متوسط | کم |
مکانیک انقباض ماهیچه اسکلتی
در بخش قبلی واکنشهای مولکولی انقباض در فیبرهای ماهیچه اسکلتی را توضیح دادیم. در این بخش نیروهای ایجاد شده در انقباض ماهیچه، تغییر طول ماهیچهها در انقباض و سرعت انقباض این بافت را بررسی میکنیم.
تحریک سلولهای ماهیچه و انقباض آنها به سرعت تحریک (تعداد پیامهای عصبی در واحد زمان) و شدت تحریک (تعداد استیل کولین متصل شده به کانالهای لیگاندی سدیم) بستگی دارد. در شروع انقباض ماهیچه اسکلتی هر نورون حرکتی فیبر ماهیچهای خود را تحریک میکند. در نتیجه فیبرهای ماهیچهای جدا از هم و بسیار خفیف منقبض میشوند. به این نوع انقباض تکانه یا «توییچ» (Twitch) گفته میشود. بر اساس نوع فیبر ماهیچهای، تکانه ممکن است از ۱۰ تا ۱۰۰ میلی ثانیه طول بکشد. نیروی ایجاد شده در هر انقباض تکانهای را میتوان در واحد زمان و بهوسیله میوگرام اندازهگیری کرد. نیروی ایجاد شده در هر تکانه ماهیچه اسکلتی را در سه بخش بررسی میکنیم.
- مرحله خاموش: در این مرحله اکتین و میوزین ماهیچه اسکلتی به هم متصل میشوند. اما حرکت نمیکنند. در نتیجه نیرویی ایجاد نمیشود.
- مرحله انقباض: میوزینهای متصل به اکتین این رشتههای پروتئینی را به مرکز سارکومر و خط M نزدیک میکنند. در این مرحله فاصله بین دو خط Z کاهش مییابد و نیروی کششی ایجاد میشود.
- مرحله استراحت: در این مرحله یون کلسیم از سارکوپلاسم خارج و میوزین از اکتین جدا میشود. فاصله بین دو خط Z به حالت قبل برمیگردد و تولید نیروی کششی متوقف میشود.

انقباض تکانهای منجر به ایجاد پتانسیل عمل و حرکت ماهیچه نمیشود. برای شروع حرکت نیاز است سارکولمای ماهیچه اسکلتی چند بار و با فاصله زمانی کم تحریک شود تا پتانسیل الکتریکی و پاسخ پایه ماهیچه (Graded Muscle Response) ایجاد شده در سلول به پتانسیل آستانه تحریک برسد. اگر قبل از تمام شدن تکانه اول سلول ماهیچهای دوباره تحریک شود، غلظت کلسیم در سارکوپلاسم بیشتر از تکانه قبلی است. به همین دلیل تکانه دوم شدیدتر خواهد بود. در واقع اثر دو تحریک با هم جمع شده و «موج تجمعی» (Wave Summation) ایجاد میکنند.
اگر تعداد تحریکها افزایش یابد، پتانسیل سلول به آستانه تحریک و نیروی کششی ایجاد شده به حداکثر میرسد. در نتیجه ماهیچه منقبض میشود. به این شرایط که فاصله بین دو مرحله استراحت تکانه ماهیچه کاهش مییابد، تتانی ناقص گفته میشود. بسیاری از کارهای روزانه ما بهوسیله این نوع انقباض انجام میشود. اگر سرعت تحریک به قدری زیاد باشد که فیبر ماهیچهای قبل از ورود به مرحله استراحت دوباره تحریک شود، تتانی کامل ایجاد میشود. این نوع انقباض به جابهجا کردن اجسام بسیار سنگین کمک میکند.

انواع انقباض ماهیچه اسکلتی
برای درک بهتر انواع انقباض ماهیچه اسکلتی خود را حین وزنه زدن در باشگاه ورزشی تصور کنید. اگر وزن وزنه زیاد باشد، شما برای برداشت وزنه از زمین تلاش میکنید اما تغییر در مکان وزنه و موقعیت دست شما ایجاد نمیشود. در این شرایط ماهیچههای اسکلتی شما منقبض میشوند اما طول انها تغییر نمی کند تا بتوانید وزنه را بلند کنید. از طرفی نیروی ایجاد شده بر اثر انقباض ماهیچه برای غلبه بر نیروی گرانش کافی نیست. به این نوع انقباض ماهیچه اسکلتی، انقباض ایزومتریک یا بدون تغییر طول ماهیچه میگویند.

اما اگر نیروی ایجاد شده از انقباض ماهیچه شما برای غلبه بر نیروی گرانش کافی باشد، میتوانید وزنه را با انقباض ماهیچه بالا و پایین حرکت دهید. با هر بار بالا بردن وزنه طول ماهیچه اسکلتی متصل به استخوان بازو و ساعد کاهش (انقباض کانسنتریک) و با هر بار پایین بردن آن طول این ماهیچه افزایش مییابد (انقباض آسنتریک). به این نوع انقباض ماهیچه اسکلتی، انقباض ایزوتونیک یا بدون تغییر نیروی انقباضی میگویند.

تونوس ماهیچه اسکلتی
ماهیچه اسکلتی به ندرت در استراحت کامل قرار دارد. حتی در زمانی که حرکت نمیکنیم، فیبرهای ماهیچه اسکلتی در بافتهای مختلف برای ثابت نگهداشتن موقعیت بدن، استخوانها و غضروفها منقبض میشوند. به این انقباض تونوس ماهیچه اسکلتی گفته میشود. برای ایجاد تونوس ماهیچهای، فیبرهای عصبی در هر زمان واحد حرکتی متفاوتی در بافت را تحریک میکنند. به این ترتیب انقباض یک دسته فیبر با استراحت دسته دیگر همراه است.
خستگی ماهیچه اسکلتی
خستگی ماهیچه اسکلتی به شرایطی گفته میشود که قدرت انقباضی ماهیچه کاهش مییابد. این حالت را به دو گروه خستگی مرکزی و محیطی تقسیم میکنند. خستگی مرکزی یا خستگی روانی مربوط به احساس ناخوشایندی است که بر اثر خستگی در فرد ایجاد میشود. این خستگی به دلیل ارسال پیام ترکیبات شبیمیایی ایجاد شده در انقباض ماهیچه به مغز ایجاد میشود. خستگی روانی، خستگی محیطی را به دنبال دارد و کمی قبل از زمانی ایجاد میشود که فیبرهای ماهیچه دیگر قادر به انقباض نباشند.
خستگی محیطی در سیناپس عصب-عضله یا در فیبرهای ماهیچهای اجاد میشود. این نوع خستگی را میتوان به دو گروه کمتکرار (Low Frequency | زمان دوی ماراتن) یا پرتکرار (High Frequency | تمرینات ورزشی سنگین) تقسیم کرد. خستگی پرتکرار به دلیل بههم خوردن تعادل یونها در غشای پلاسمایی ایجاد میشود. در این وضعیت پمپ سدیم-پتاسیم کارایی کافی ندارد. در نتیجه کانالهای ولتاژی سدیم و کلسیم نمیتوانند اختلاف پتانسیل مناسب و پیام عصبی در سلول ایجاد کنند. خستگی پرتکرار ماهیچههای اسکلتی معمولا پس از ۳۰ دقیقه بر طرف میشود. خستگی کمتکرار به دلیل اختلال در آزاد شدن یون کلسیم ایجاد میشود. یرطرف شدن خستکی کمتکرار به زمان بیشتری (از ۲۴ تا ۷۲ ساعت) نیاز دارد.
تجمع فسفات معدنی در سارکوپلاسم، کاهش pH بافت به دلیل افزایش یون هیدروژن، تمام شدن ذخیره گلیکوژن بافت و بهمریختن تعادل غلظت پتاسیم بین سارکوپلاسم و ماتریکس خارج سلولی، از عوامل دیگری است که بر ایجاد خستگی ماهیچه اثر دارد.
حرکت ماهیچه اسکلتی
برای حرکت دادن هر یک از اندامهای بدن معمولا یک جفت ماهیچه اسکلتی وجود دارد. اتصال این ماهیچهها به استخوانهای بدن، سرعت، نیروی ایجاد شده و میزان حرکت اندام را مشخص میکند. مجموعه استخوان و ماهیچه متصل به آن شبیه یک اهرم کار میکند. اهرم از یک جسم میلهای محکم تشکیل شده است که دور یک نقطه اتکای ثابت حرکت میکند. شکل زیر یک اهرم ساده را نشان میدهد.

در شکل بالا نیروی گشتاور، نیرویی است که شما برای بلند کردن جسم به میله وارد میکنید. فاصله نقطه وارد شدن این نیرو تا نقطه اتکا بازوی نیرو نام دارد. نیروی مقاومتی، نیرویی است که به دلیل وزن جسم در برابر بالا آمدن مقاومت میکند. فاصله این نیرو تا نقطه اتکا بازوی مقاومتی نام دارد. اگر نیرویی که شما به میله وارد میکنید از نیروی مقاومتی جسم بیشتر باشد، سنگ از زمین بلند میشود. اگر بازوی نیرو از بازوی مقاومت بلندتر باشد، نیروی بیشتری ایجاد میشود. به کمک این سیستم میتوان اجسام بسیار سنگین را در فاصله کمی جابهجا کرد. جک ماشین به این شیوه حرکت میکند. اما اگر بازوی مقاومت بلندتر از بازوی نیرو باشد، نیروی کمتری ایجاد میشود که میتواند جسم سبکتری را با سرعت در فاصله بیشتر جابهجا کند. ضربه چوب بیسبال به توپ نمونهای از این اهرم است.
در بدن انسان استخوان بازوی سخت اهرم، مفصل نقطه اتکا و ماهیچه منبع نیرویی است که برای جابهجا کردن اجسام وارد میشود. بازوی نیرو در این سیستم فاصله بین مفصل (نقطه اتکا) تا انتهای ماهیچه (نیروی گشتاور) و بازوی مقاومتی فاصله بین مفصل تا مرکز ماهیچه (جسم) است. اهرمهای بدن بر اساس جایگاه مفصل، ماهیچه و استخوان به سه دسته تقسیم میشوند.
اهرم نوع اول
مفصل بین بازوی نیرو و مقاومت قرار دارد. بر اساس فاصله مفصل تا مرکز ماهیچه مجموعه استخوان-ماهیچه مقاومتی یا سرعتی عمل میکند. زمانی که ماهیچه پشت گردن سر شما را بالا میآورد، مجموعه ماهیچه-استخوان یک اهرم نوع اول است. ماهیچههای اسکلتی پشت گردن نیروی لازم برای بلند کردن سر (جسم) را فراهم میکند و مفصل بین سر و گردن نقطه اتکا است.
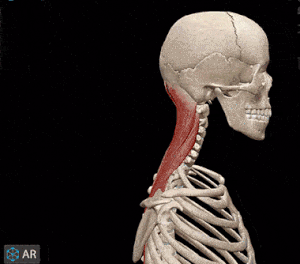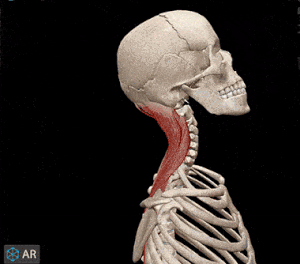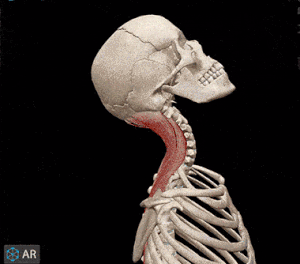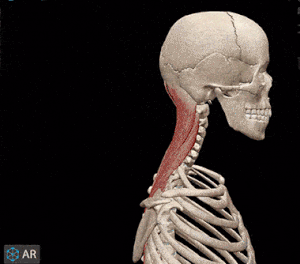
اهرم نوع دوم
در این اهرم جسم بین انتهای ماهیچه (محل وارد شدن نیرو) و مفصل قرار میگیرد. برای درک این حرکت در بدن تصور کنید روی صندلی نشستهاید و زانوهای شما با زاویه ۹۰ درجه قرار دارند. در این وضعیت برای بلند کردن پاشنه پا از روی زمین بدن شما از اهرم نوع دو کمک میگیرد و شما میتوانید تمام وزن پا را با ایجاد نیروی کم، در فاصلهای کم رو به بالا حرکت دهید (اهرم قدرتی).

اهرم نوع سوم
در این نوع اهرم نیروی منتقل شده از انتهای ماهیچه به استخوان بین مفصل و جسم قرار دارد. خم کردن ساعد به کمک انقباض ماهیچه دوسر بازو مثالی از این اهرم سرعتی در بدن است. بیشتر ماهیچههای اسکلتی و استخوانهای متصل به آنها در بدن انسان از این اهرم برای حرکت بهره میبرند. میتوان گفت بدن انسان برای جابهکردن اجسام سبک (بخشهای بدن) با سرعت و در فاصلهای طولانی طراحی شده است. به همین دلیل در شرایط خطر میتواند خیلی سریع فرار کند.

آناتومی ماهیچه اسکلتی
در بدن هر فرد بالغ بیش از ۶۰۰ ماهیچه اسکلتی وجود دارد که اندازه و آرایش فیبرهای ماهیچهای در آنها بسیار متفاوت است. آِرایش دستههای فیبر ماهیچهها یا فاسیکولها شکل نهایی بافت ماهیچهای، جهت حرکت استخوان پس از انقباض ماهیچه و میزان نیروی تولید شده از انقباض را تعیین میکند. آرایش موازی، حلقه و پَر سه تقسیمبندی متداول برای آرایش فاسیکولهای ماهیچه اسکلتی است.
- آرایش موازی: در این آرایش فاسیکولهای ماهیچه اسکلتی از طول کنار هم قرار میگیرند. این آرایش در شکلدهی بیشتر ماهیچههای اسکلتی بدن نقش دارد. آرایش موازی فاسیکولها در بعضی ماهیچههای اسکلتی ازجمله «ماهیچه خیاطه»(Sartorius) صفحههای صاف و بلندی تشکیل میدهد که سطح زیادی برای اتصال به استخوان دارند. اما این آرایش در بعضی از بافتهای ماهیچهای مثل ماهیچههای دوسر بازو ساختاری سبیه دوک تشکیل میدهد و فقط دو سر انتهایی ماهیچه بهوسیله زردپیها به استخوان متصل میشوند.
- آرایش حلقوی: ماهیچههای حلقوی یا اسفنکترها اطراف دهان، مردمک چشم، انتهای روده بزرگ و میزراه قرار دارند. فیبرهای ماهیچهای در این آرایش شبیه یک حلقه متراکم کنار هم قرار میگیرند. استراحت ماهیچههای اسکلتی حلقوی با باز شدن دریچههای بدن و انقباض آنها با بسته شدن دریچهها همراه است. برای مثال در صوت زدن ماهیچههای حلقوی اطراف دهان منقبض و لبهای شما جمع میشود.
- آرایش مورب یا همگرا: در آرایش مورب یا همگرا فاسیکولهای ماهیچه اسکلتی در یک نقطه به هم میرسند.نقطه اتصال این فیبرها یک رشته تاندون، صفحهای از بافت پیوندی یا آپونوروزیس یا زردپی بسیار باریک و کوچک است. فیبرهای ماهیچهای در این آرایش منطقه وسیعی از بدن را به خود اختصاص میدهند. ماهیچههای قفسه سینه (ماهیچه پکتوریال) یکی از ماهیچههای موربی است که بهوسیله یک تاندون به قسمت بالایی استخوان بازو متصل میشود.
- آرایش پَر یا «پنات» (Pennate): در این آرایش فیبرهای ماهیچه اسکلتی دور یک تاندون مرکزی میچرخند. تاندون در این آرایش شبیه نی میانی پر پرندگان و فیبرهای ماهیچه اسکلتی شبیه زائدههای خروجی از آن هستند. در این آرایش فیبرهای ماهیچهای فقط در یک زاویه منقبض میشوند و تاندون متصل به آنها حرکت زیادی ندارد. اما ازآنجایی که تعداد فیبرهای ماهیچهای نسبت به اندازه کل ماهیچه در این آرایش بیشتر است، قدرت انقباضی ماهیچههایی که این ساختار را دارند، بیشتر است. فیبرهای ماهیچهای با سه آرایش به تاندون متصل میشوند.
- آرایش تکپر: در این آرایش فاسیکولها در یک جهت تاندون میانی قرار دارند. ماهیچه راستگرد ساعد با این آرایش تشکیل میشود.
- آرایش دوپر: در این آرایش فاسیکولها در دو طرف تاندون میانی (ساختار اصلی پر پرنده) قرار میگیرند.
- آرایش چندپر: در این آرایش فاسیکولها در دو طرف تاندونهای مختلف قرار دارند. این تاندونها به هم متصل میشوند و تاندون اصلی ماهیبچه را تشکیل میدهند. آرایش فیبرهای ماهیچهای در بافت ماهیچه دلتای شانه از این نوع است.

ماهیچه اسکلتی در کدام بخش های بدن قرار دارد ؟
کوچکترین ماهیچههای اسکلتی در گوش میانی به استخوانچههای شنوایی متصل هستند و بزرگترین بافت ماهیچه اسکلتی در ران قرار دارد. بخشی از ماهیچه اسکلتی که به استخوان ثابت متصل میشود، «مبدا» (Origin) ماهیچه و بخشی از آن که به استخوان متحرک متصل میشود، نقطه اتصال یا «اینزرشن» (Insertion) نام دارد. بافت ماهیچه اسکلتی بدن را میتوان در چهار بخش سر و گردن، تنه، اندامهای حرکتی بالایی (دستها) و اندامهای حرکتی پایینی (پاها) بررسی کرد.
- ماهیچههای سر و گردن: این ماهیچه شامل ماهیچههای حالت چهره، ماهیچههای جویدن، ماهیچههای خارج چشمی، ماهیچه زبان، ماهیچه حلق، ماهیچه حنجره و ماهیچههای گردن میشود.
- ماهیچههای تنه: این ماهیچهها شامل ماهیچههای پشت، ماهیچههای بخش جلویی و کناری ناحیه شکمی و ماهیچههای لگن میشود.
- ماهیچههای اندام حرکتی بالایی: این ماهیچهها شامل ماهیچههای شانه، بازو، ساعد و کف دست میشوند.
- ماهیچههای اندام حرکتی پایینی: این ماهیچهها شمال ماهیچههای پهلو، ران، ساق و کف پا میشوند.
ماهیچه های اسکلتی سر و گردن
حدود ۲۰ بافت ماهیچه اسکلتی صاف زیر پوست صورت و جمجمه قرار دارد. بر خلاف بافت ماهیچه اسکلتی بخشهای دیگر بدن، غلاف بافت پیوندی خارجی (فاسیکا) روی این ماهیچهها قرار ندارد. ماهیچههای صورت کنار ورودی چشم، بینی، گوش و دهان قرار دارند. به همین دلیل آنها به ماهیچههای دهانی (گروه بوکولابیال | Buccolabial Group)، ماهیچههای بینی (گروه نازال | Nasal Group)، ماهیچههای جمجمه و گردن (گروه اپیکارنیال | Epicranial Group)، ماهیچههای گوش خارجی (گروه اوریکولار | Auricular Group) و ماهیچههای پلک (گروه اوربیتال | Orbital Group) تقسیم میکنند. این ماهیچهها در ایجاد حالات چهره ازجمله لبخند و اخم نقش دارند. پیام عصبی سیستم عصبی مرکزی بهوسیله اعصاب جمجمهای ۷ (CN IIV) به این بافتهای ماهیچهای منتقل میشود.

ماهیچههای جویدن مجموعهای بافتهای ماهیچهای گیجگاهی، فک، پتریگوئید میانی و کناری هستند. این ماهیچهها به حرکت جهات بالا، پایین، چپ و راست فک پایینی برای جویدن لقمه غذا کمک میکنند. پیام عصبی سیتم عصبی مرکزی بهوسیله اعصاب جمجمهای ۵-۳ (CN V3) به این باتهای ماهیچه اسکلتی منتقل میشود.

در هر چشم، ۶ ماهیچه اسکلتی کره چشم را به استخوان حفره چشم (استخوان اوربیت) متصل میکند. ۴ ماهیچه کره چشم را در جهات جغرافیایی شمال، جنوب، شرق و غرب حرکت میدهند و ۲ ماهیچه اسکلتی حرکت چشم را با حرکت سر هماهنگ میکنند. پیام سیستم عصبی مرکزی بهوسیله عصب جمجمهای ۳ (CN II | عصب حرکتی-چشمی)، عصب جمجمهای ۴ (CN IV |عصب قرقرهای ) و عصب جمجمهای ۶ (CN VI | عصب دورکننده) به ماهیچههای خارج چشمی منتقل میشود.

ماهیچه زبان بخشی از بافت ماهیچه بدن در سیستم گوارش است. این ماهیچه به درک مزه، جویدن لقمه، بلع غذا، تکلم و پاکسازی حفره دهان کمک میکند. به همین دلیل اعصاب حرکتی و حسی فراوانی با سلولهای این بافت ماهیچهای سیناپس دارند. اعصاب جمجمهای ۱۲ (CN XII | عصب زیرزبانی)، عصب جمجمهای ۷ (CN VII | عصب صورت)، عصب جمجمهای ۹ (CN IX | عصب زبانی-حلقی) و عصب جمجمهای ۱۰ (CN X | عصب واگ) پیام عصبی سیستم عصبی مرکزی را به این ماهیچه یا تحریک گیرندههای چشایی را به سیستم عصبی مرکزی منتقل میکنند. ماهیچه زبان تنها ماهیچه اسکلتی است که به استخوان متصل نمیشود.
وظیفه اصلی ماهیچههای گردن حفظ موقعیت سر و کمک به حرکات آن در جهات مختلف است. این ماهیچهها به سه گروه اصلی جلویی (حرکت سر به جلو)، کناری (حرکت سر به چپ و راست) و پشتی (حرکت سر به عقب) تقسیم میشوند. هر یک از این گروهها را میتوان بر اساس عمق بدن، عملکرد و جایگاه دقیق به گروههای تخصصیتر تقسیم کرد.

ماهیچه های اسکلتی تنه
ماهیچههای اسکلتی تنه در سه بخش پشت، ناحیه شکمی و کف لگن به حفظ ساختار بدن، حفاظت اندامهای ناحیه شکمی و حرکت لگن کمک میکنند. ماهیچههای اسکلتی پشت را میتوان در دو گروه اصلی بررسی کرد.
- ماهیچههای خارجی یا سطحی: این ماهیچهها به حرکت اندامهای فوقان، حفظ موقعیت بدن، حرکت گردن، بالا کشیدن قفسه سینه در تنفس کمک میکنند و به دو دسته سطحی و میانی تقسیم میشوند.
- ماهیچههای درونی یا عمقی: وظیفه اصلی این ماهیچهها کمک به حرکت ستون مهرهها است. این ماهیچهها به سه دسته سطحی، عمقی و عمیقتر تقسیم میشوند.

ماهیچههای اسکلتی ناحیه شکمی زیر پرده صفاق قرار دارد و مثل یک تونل بافت ماهیچهای از اندامهای گوارشی محافظت میکند. در تنفس عمیق، انقباض ماهیچههای شکمی جلویی به کاهش حجم قفسه سینه کمک میکند و برای چرخش تنه، ماهیچههای کناری این بخش منقبض میشوند.

ماهیچه اسکلتی لگن
ماهیچههای اسکلتی لگن به بخشهای انتهایی استخوان لگن متصل میشوند و بخشی از آرایش آنها اسفنکترهای خارجی میزراه و مقعد را ایجاد میکند. به علاوه این ماهیچهها کمک میکنند استخوان لگن در موقعیت خود ثابت بماند و بر نیروی گرانش وارد شده بر آن غلبه کند. ماهیچههای اسکلتی لگن در زنان، به تولد نوزاد کمک میکند.

ماهیچه های اسکلتی اندام حرکتی فوقانی
انقباض ماهیچههای اسکلتی شانه، بازو، ساعد و کف دست به حرکت اندام فوقانی در جهات مختلف کمک میکند.
بازو بخشی بین شانه تا آرنج است که از سه ماهیچه جلویی (ماهیچه دوسر، ماهیچه بازویی و ماهیچه کوراکوئید) و یک ماهیچه پشتی (ماهیچه سهسر بازویی) تشکیل میشود. انقباض ماهیچههای دوسر و سهسر بازویی ساعد را خلاف جهت هم حرکت میدهد. انقباض ماهیچه دوسر بازویی با کاهش زاویه بازو و ساعد و بالا آمدن ساعد همراه است. انقباض ماهیچه سهسر بازو با افزایش زاویه بین بازو و ساعد و پایین آمدن ساعد همراه است. نامگذاری این ماهیچههای اسکلتی به دلیل تعداد انتهای اتصالی به استخوان ثابت است.
ماهیچه اسکلتی اندام حرکتی تحتانی
ماهیچههای اسکلتی اندام تحتانی به استخوانهای ران، زانو، ساق، مچ و کف پا متصل میشوند. بخش بالایی این ماهیچهها به استخوان ثابت لگن و انتهای دیگر آنها به استخوان فمور متصل میشود. بیشتر وزن ماهیچهای این بخش از بدن مربوط به ماهیچههای پشتی است که وظیفه حرکت ساق را برعهده دارند. ماهیچههایی که در ساق پا قرار دارند به سه دسته جلویی، پشتی و کناری تقسیم میشوند. ماهیچه خیاطه، ماهیچه راست فمور، ماهیچه پهن جانبی و میان، ماهیچه گاستروکیمونس، ماهیچه درشتنی و ماهیچه نعلی، هفت ماهیچه اسکلتی در بخش پشتی پا هستند که انقباض آنها پا را به جلو و عقب حرکت میدهد. عضله چهار سر به بخش جلویی استخوان فمور در ران متصل میشود. انقباض این ماهیچه سبب صاف شدن پا از محل زانو میشود. انقباض ماهیچه درشتنی پشتی و جلویی به حرکت (بالا و پایین) مچ پا کمک میکند.

انعکاس ماهیچه اسکلتی
پاسخهای انعکاسی به فعالیتهای غیرارادی گفته میشود که در پاسخ به محرکهای محیطی و بهوسیله عصبهای نخاع کنترل میشوند. انعکاس ماهیچه اسکلتی یکی از متدوالترین فعالیتهای غیرارادی بدن است. انعکاس کشش ماهیچه اسکلتی یکی از متدوالترین فعالیتهای انعکاسی بدن است. مسیر عصبی این انعکاس از گیرندههای حس موقعیت در ماهیچههای اسکلتی، نورونهای حسی آوران، مرکز تجمع پیام عصبی (نخاع)، نورون حرکتی وابران و ماهیچه اسکلتی تشکیل شده است.
ماهیچههای اسکلتی از دو دسته فیبر مختلف تشکیل شده است. فیبرهای خارجی با تاندونها در ارتباط هستند و نیروی حاصل از انقباض را به استخوانها منتقل میکنند. این بخش از ماهیچه فیبرهای مخططی هستند که فیبرهای خارجی نام دارند.این ماهیچهها مسئول انقباض و ایجاد حرکت استخوانها هستند. زیر این فیبرها، کپسولی از بافت پیوندی اطراف فیبرهای دوک داخلی قرار دارد. فیبرهای داخلی در حرکت نقشی ندارند اما حرکت ماهیچه و موقعیت آن در فضا را شناسایی میکنند. این فیبرها گیرندههای حس موقعیت در ماهیچه هستند. این گیرندهها تغییر طول ماهیچه و سرعت حرکت آن را تشخیص میدهند.

فیبرهای داخلی مثل فیبرهای انقباضی ماهیچههای اسکلتی سلولهایی با طول زیاد و چند هستهای هستند. در هر بافت ماهیچه اسکلتی دو نوع فیبر داخلی یا گیرنده حس موقعیت قرار دارد که بر اساس محل قرار گیری هسته به دو نوع هسته زنجیرهای (هستهها متوالی در طول سلول) و هسته کیسهای (تجمع هستهها در مرکز سلول) تقسیم میشوند. گیرندههای هسته کیسهای به محرکهای تغییر طول ماهیچه و سرعت تغییر حساستر هستند و گیرندههای هسته کیسهای به محرکهای تغییر طول و شدت کشش حساستر هستند.
دورتادور هستههای هر فیبر داخلی پایانه نورونهای حسی نوع I (یا انتهای فنری نورون حسی) قرار دارد. تغییر طول فیبر داخلی و کشش آن با باز شدن دریچه کانالهای مکانیکی سدیم در غشای این فیبرهای عصبی، تغییر پتانسیل غشا، باز شدن کانالهای سدیم ولتاژی ایجاد پیام عصبی و انتقال آن به نخاع همراه است. فیبرهای داخلی زنجیرهای علاوه بر نورونهای حسی نوع I، با نورونهای حسی نوع II سیناپس دارند. دو طرف هستههای فیلامنتهای اکتین و میوزینی قرار دارد به استیلکولین آزاد شده از نورونهای حرکتی گاما پاسخ میدهند و حرکت آنها منجر به کشش فیبرهای داخلی میشود.
برای درک بهتر عملکرد این گیرندهها تصور کنید تاندون متصل به ماهیچه چهارسر با ضربه تحریک میشود. کشش ایجاد شده از ضربه در تاندون، سبب کشش فیبرهای ماهیچهای خارجی و گیرندههای حس موقعیت خواهد شد. پیام عصبی ایجاد شده بهوسیله نورونهای حسی نوع I و II از شاخه پشتی وارد نخاع میشود. این نورونها با نورونهای متفاوتی در نخاع سیناپس تشکیل میدهند.
- سیناپس این نورون با نورونهای حرکتی آلفا پیام عصبی را از ریشه شکمی نخاع به فیبرهای خارجی ماهیچههای چهارسر ران منتقل میکند. انقباض ماهیچه چهارسر با صاف شدن پا در محل زانو همراه است.
- سیناپس پایانه آکسون نورون حسی با نورونهای بینابینی، مسیر انتقال پیام از نورونهای حرکتی آلفا به فیبرهای خارجی ماهیچههای پشت ران و نورونهای گامای فیبر خارجی ماهیچه پشت ران را مهار میکند.
- سیناپس این نورونهای حسی با نورونهای حرکتی گاما، پیام عصبی نخاع را به گیرنده حس موقعیت ماهیچه چهارسر منتقل و این گیرندهها را تحریک میکند تا کشش زیاد و آسیب ماهیچه اسکلتی جلوگیری کند.

رفلاکس ماهیچه دوسر بازو یکی از فعالیتهای غیرارادی بدن است که دست شما را از آسیب دیدن حفظ میکند. تا به حال توجه کردهاید، زمانی که جسم بسیار داغی را لمس میکنید بازوی شما ناخودآگاه و سریع جمع میشود. این واکنش یک انعکاس نخاع-ماهیچه اسکلتی است. در این وضعیت تحریک گیرندههای دمایی پوست دست بر اثر تغییر دما تحریک شده و پیام آنها به نخاع منتقل میشود. در نخاع نورون حسی پبا نورون حرکتی آلفا سیناپس تشکیل میدهد و انتقال پیام عصبی به فیبرهای خارجی ماهیچه دوسر با انقباض ماهیچه و جمع شدن دست همراه است.
بیماری های ماهیچه اسکلتی
در بخشهای قبلی توضیح دادیم که انقباض ماهیچه اسکلتی به فعالیت هماهنگ سیستم عصبی، کانالهای غشایی و پروتئینهای اسکلت سلولی این ماهیچهها وابسته است. پس هر عامل فیزیلوژیک یا پاتولوژیکی که در فعالیت این سه عامل اختلال ایجاد کند با بیماری ماهیچههای اسکلتی همراه است. دیستروفی دوشن و دیستروفی بکر دو اختلال ایجاد شده بر اثر جهش پروتئینهای فیبر ماهیچه اسکلتی هستند.
- دیستروفی دوشن: این اختلال یک بیماری ژنتیکی وابسته به X و مغلوب و به همین دلیل احتمال ابتلا به آن در مردها بیشتر از زنان است. علائم این بیماری در سن ۵ تا ۶ سالگی ایجاد میشود. در این اختلال پروتئین دیستروفین سنتز نمیشود. در نتیجه به مرور زمان سارکوپلاسم از بین میرود و فیبر ماهیچهای به بافت فیبروزی و چربی تبدیل میشود. دیافراگم یکی از ماهیچههای اسکلتی مهم در بدن است که تحلیل رفتن در دیستروفی دوشن منجر به اختلال در تنفس میشود.
- دیستروفی بکر: در این اختلال جهش ژنتیکی کدونها منجر به تغییر در توالی آمینواسیدی و تاخوردگی پروتئین میشود. علائم این بیماری معمولا در سن ۱۰ تا ۱۱ سالگی ایجاد میشود.
بیماری میاستنی گراویس یکی از بیماریهای خودایمنی دستگاه حرکتی انسان است. در این بیماری بدن گیرندههای نیکوتینی (کانالهای لیگاندی سدیم) ماهیچه اسکلتی را پاتوژن شناسایی و علیه آن آنتیبادی تولید میکند. در نتیجه استیل کولین نمیتواند پیام عصبی نورون حرکتی را به فیبر ماهیچهای منتقل کند. این بیماری پیشرفت سریعی دارد و منجر به فلج فرد میشود. سندروم ایتن لامبرت یکی دیگر از بیماریهای خودایمنی است که دستگاه حرکتی انسان را مختل میکند. در این بیماری سیستم ایمنی کانالهای ولتاژی کلسیم در پایانه آکسون در نورونهای حرکتی را پاتوژن شناسایی و علیه آن آنتیبادی تولید میکند. در نتیجه جریان کلسیم به پایانه آکسون، اگزوسیتوز وزیکولهای سیناپسی و اتصال استیل کولین به گیرندههای سارکولما مهار میشود. علائم این بیماری شبیه بیماری میاسنتی گراویس است. مهارکنندههای آنزیم استیل کولین ترانسفراز یکی از داروهایی است که برای جلوگیری از پیشرفت این بیماری تجویز میشود. با مهار این آنزیم غلظت استیل کولین موجود در شکاف سیناپسی و تحریک ماهیچه افزایش مییابد.
توکسین بعضی باکتریها (بوتولیوم و تتانوس) با مهار پروتئینهای اگزوسیتوزی (سیناپتوتاکسین و سیناپتوبوین) در پایانه آکسون نورون حرکتی از اتصال وزیکول سیناپسی به غشای نورون و آزاد شدن انتقال دهنده عصبی به شکاف سیناپسی و تحریک ماهیچه جلوگیری میکند. در نتیجه ماهیچه فلج میشود. دندروتاکسین سم ماری است که به کانالهای پتاسیمی پایانه آکسون متصل شده و خروج یون پتاسیم از سلول و ریپلاریزاسون غشا را مهار میکند. در نتیجه غشا ورود کلسیم به سلول و اگزوسیتوز وزیکولهای استیل کولین ادامه مییابد. بانگروتاکسین آلفا (سم مار کبری) یکی دیگر از مولکولهای شیمیایی است که عملکرد کانالهای ساکولما را تغییر میدهد. این سم کانالهای لیگاندی سدیم و تحریک سلول ماهیچه اسکلتی را مهار میکند.
سوالات متدوال
در این بخش به تعدادی از سوالات متداول پیرامون ماهیچه اسکلتی پاسخ میدهیم.
تفاوت ماهیچه اسکلتی و قلبی چیست ؟
ماهیچه قلبی و اسکلتی دو بافت مهم بدن هستند که مهمترین تفاوت آنها در عامل تحریک انقباض است. ماهیچههای اسکلتی به شکل ارادی و با پیامهای قشر مخ منقبض میشوند. اما انقباض ماهیچه قلب یک فعالیت غیرارادی است که بهوسیله سیستم خودتحریکی قلب شروع میشود. فیبرهای ماهیچه قلبی معمولا از دو تا سه هسته تشکیل میشوند و منشعب هستند. بین هر دو فیبر ماهیچه قلبی کنار هم، صفحهای از جنش غشای پلاسمایی قرار دارد که از ادغام غشای فیبرهای تشکل شده است.

متابولیسم آنها برخلاف سلولهای ماهیچهای به اکسيژن وابسته است. مولکول ATP لازم برای انقباض این ماهیچهها از مسیرهای هوازی (گلیکولیز و زنجیره انتثال الکترون) در میتوکندری تولید میشود. مویرگهای خونی و میوگلوبینهای فراوان اکسیبژن لازم برای تولید ATP را تامین میکنند. فیبرهای پروتئینی این این ماهیچه شبیه فیبرهای پروتئینی ماهیچه اسکلتی است. به همین دلیل شکل ماهیچه قلبی زیر میکروسکوپ با خطوط تیره و روشن دیده میشود. این ماهیچه فقط در قلب وجود دارند.
تفاوت ماهیچه اسکلتی و صاف چیست ؟
ماهیچه صاف یکی از بافتهای مهم بدن در دستگاه گوارش و تفاوت اصلی آن با ماهیچه اسکلتی شیوه انقباض است. در ماهیچه اسکلتی کلسیم آزاد شده از شبکه سارکوپلاسمی با اتصال به تروپونین، اتصال اکتین و میوزین را تسهیل میکند. اما این یون در ماهیچه صاف به پروتئین کلمودولین متصل میشود. به علاوه پتانسیل الکتریکی در ماهیچه اسکلتی فقط بر اثر اتصال انتقالدهنده عصبی و گیرندههای سارکولما ایجاد میشود. اما در ماهیچه صاف علاوه بر تحریک نورونهای عصبی، انتقال کلسیم از سلولهای ... به ماهیچه با ایجاد پتانسیل نیزهای همراه است.

ماهیچههای اسکلتی بافتی متصل به استخوانها هستند که به حرکت بدن در جهات مختلف کمک میکنند. اما ماهیچههای صاف دیواره مجاری بدن را میسازند. بیشتر ماهیچههای صاف بدن در لوله گوارش قرار دارد و انقباض غیرارادی ان به گوارش فیزیکی غذا و حرکت آن در این لوله کمک میکند. هر فیبر ماهیچه صاف از یک هسته تشکیل شده است و آرایش رشتههای اکتین و میوزین در این سلولها مثل سلولهای ماهیچه قبلی و اسکلتی در به شکل خوط موازی و خطی نیست. در این سلولها اکتین و میوزین در واحدهای پلکانی قرا میگیرند. به همین دلیل خطهای تیره و روشن در تصویر میکروسکوپی این ماهیچه وجود ندارد.
نوار عصب غضله چیست ؟
الکترومیوگرافی (Electromyography | EMG) یا نوار عصب-عضله یکی از روشهای ارزیابی سلامت ماهیچهها و تحریکپذیری آنها نسبت به نورونهای حرکتی است. بهوسیله این روش میتوان اختلال نورونهای حرکتی، بافت ماهیچهای یا سیناپس ماهیچه-عصب را بررسی کرد. الکترودهای سوزنی این دستگاه وارد پوست و ماهیچه میشود. سپس جریان الکتریکی ایجاد شده در ماهیچه به سیستم کامپیوتری منتقل و به اطلاعات عددی قابل اندازهگیری تبدیل میشود.











دور هر یاخته ماهیچه غلاف پیوندی یا بافت پیوندی وجود داره؟
سلام و وقت به خیر؛
بله. دور هر سلول ماهیچه (میوسیت) شاهد حضور غلافی از جنس بافت پیوندی هستیم که به آن «اندومیوزیم» (Endomysium) گفته میشود.
از همراهی شما با مجله فرادرس سپاسگزاریم.