



کنفورماسیون مولکول های زیستی – به زبان ساده
کنفورماسیون مولکولهای زیستی، همان ساختار سهبعدی و عملکردی مولکول است که بر اثر عوامل مختلف تغییر میکند و ساختار و عملکرد جدیدی به مولکول میدهد. درک این مفهوم و چگونگی تغییر آن، اساس درک زیستشناسی مولکولی است. در این مطلب مفهوم کنفورماسیون و تاثیر تغییرات کنفورماسیونی بر عملکرد پروتئین، DNA و RNA را توضیح میدهیم.


کنفورماسیون چیست ؟
کنفورماسیون (Conformation) یا صورتبندی، سازمانیافتگی سهبعدی یک مولکول است که با چرخش حول محورهای مختلف و بدون شکستن پیوندها، تغییر میکند. کنفورماسیون مولکولهای زیستی بر اثر تغییر در عوامل محیطی ازجمله دما، غلظت یونی و pH یا اتصال لیگاند به رسپتور تغییر میکند.
تفاوت کنفورمیشن و کانفیگوریشن
هر دو مفهوم کنفورمیشن (کنفورماسیون) و «کانفیگوراسیون» (Configuration) مربوط به ساختار فضایی ترکیبات شیمیایی، بخصوص ترکیبات آلی هستند. با این تفاوت که کانفیگوراسیونهای مختلف، با شکستن پیوند در مولکول ایجاد میشوند اما در تبدیل کنفورماسیونها به هم پیوندی نمیشکند. برای نمونه ساختارهای سیس و ترانس اسیدهای چرب، کانفیگوراسیون این مولکول بهحساب میآید.
ایزومرهای کنفورماسیونی
سازمانیافتگی سهبعدی متفاوت در یک مولکول را «ایزمرهای کنفورماسیون» (Conformational Isomers) یا «کانفورمر» (Conformer) میگویند. نکته قابل توجه این است که این ایزومرها از نظر توالی و ترکیب مونومرها هیچ تفاوتی با هم ندارند و تنها جهتگیری فضایی مونومرها در آنها متفاوت است. برای نمونه دو کنفورماسیون T و S در هموگلوبین که پایینتر در مورد آن میخوانیم، دو ایزومر کنفورماسیونی هستند.
نیروهای موثر در تغییرات کنفورماسیونی
نیروهایی که به تشکیل ساختار سهبعدی مولکولهای زیستی کمک میکنند، در تغییر کنفورماسیونها نیز مشارکت دارند. در ادامه این نیروها را معرفی میکنیم.
- برهمکنشهای آبگریز: این برهمکنش بین قسمتهای ناقطبی دو مولکول یا بخشهای غیرقطبی در یک مولکول بهوجود میآيد.
- پیوندهای هیدروژنی: این پیوند از برهمکنشهای دوقطبی-دوقطبی بین دو مولکولی تشکیل میشود که در یکی از آنها، هیدروژن به اتمی با الکترونگاتیوی بالا متصل شده است. این برهمکنش نیز بین دو مولکول یا بخشهای مختلف یک ماکرومولکول بهوجود میآيد.
- نیروهای واندوالس: این برهمکنش بین دو مولکول یا بخشهای مختلف مولکولی بهوجود میآید که قطبشپذیری دارد.
کنفورماسیون پروتئین چیست ؟
کنفورماسیون پروتئین در عملکرد آن و برهمکنش این مولکول با دیگر مولکولهای آلی و معدنی نقش بسیار مهمی دارد. بر اساس تغییرات کنفورماسیونی، دو نوع برهمکنش در این مولکولها وجود دارد.
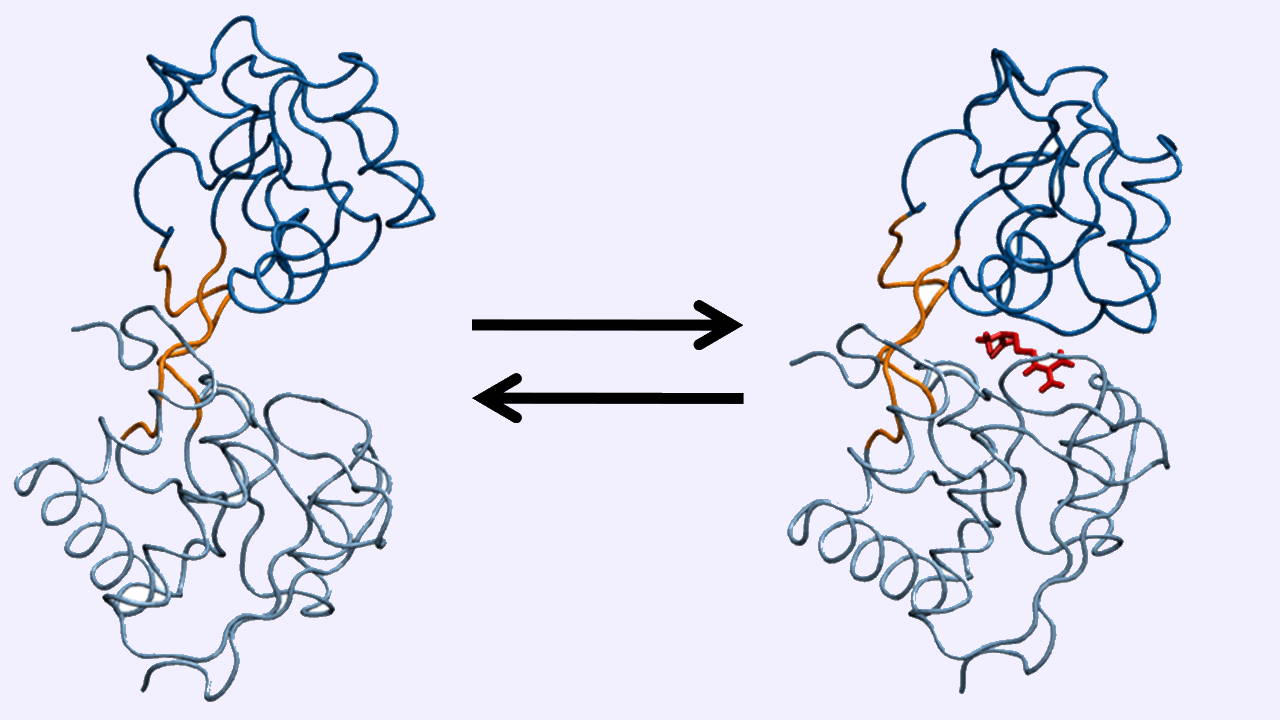
- برهمکنش «قفل و کلید» (Lock-and-Key): در این حالت پروتئین و سوبسترا، ساختاری مکمل دارند و به تغییر ساختار هیچکدام برای اتصال نیاز نیست.
- برهمکنش «جفت شدن القایی» (Induced Fit): در این حالت اتصال سوبسترا، یا عامل تنظیمی سبب ایجاد تغییر در ساختار پروتئین و جفت شدن دو مولکول میشود. این نوع برهمکنش، تنوع بیشتری دارد.
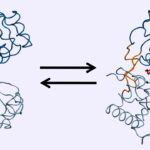
کنفورماسیون مولکول، ممکن است در سطح برهمکنش آن با مولکول دیگر یا دور از سطح تغییر کند. برای نمونه، اتصال به «همِ» (Heme) در هموگلوبین، در سطح اتصال تغییرات ایجاد میکند اما اتصال لیگاند به سطح خارج سلولی رسپتور، کنفورماسیون بخش سیتوپلاسمی آن را تغییر میدهد. نکته قابل توجه این است که تغییرات کنفورماسیونی به شکل برگشتپذیر انجام میشود و چرخهای تکراری در عملکرد سلول دارد.
این تغییرات ساختاری در سطح چهارم پروتئین، وابسته به نوع و نحوه قرارگیری زیرواحدهای آمینواسیدی در سطح اول، دوم و سوم است. برای مطالعه کامل در مورد این ساختارها میتوانید به مطلب «ساختار پروتئین در چهار سطح | به زبان ساده» مراجعه کنید.
تغییر کنفورماسیون در هموگلوبین
هموگلوبین ماکرومولکولی تترامر است و از دو زیر واحد و دو زیرواحد تشکیل میشود. هر زیرواحد بخش پروتئینی «هِم» (Heme) به متصل میشود. این پروتئین دو کنفورماسیون T (بدون اکسیژن) و R (متصل به اکسیژن) دارد. اتصال به یکی از گروههای هِم تغییرات زیر را بهدنبال دارد که تمایل هِم به را افزایش میدهند.
- تغییر در الکترونگاتیوی هیستیدین موجود در جایگاه اتصال
- تغییر برهمکنش بین زیرواحدها
- تغییر کنفورماسیون T به R
تغییر کنفورماسیون در آنزیم
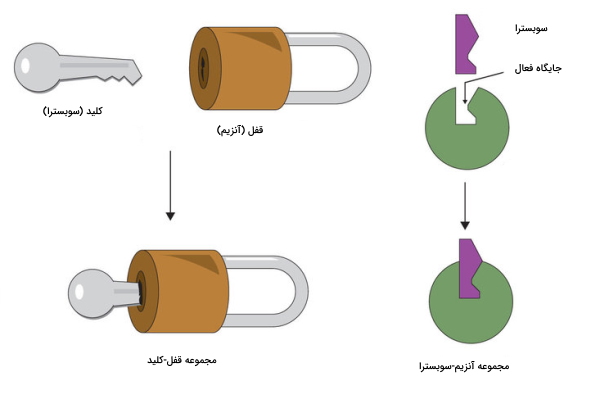
«آنزیمهای آلوستریک» (Allosteric Enzymes) ماشینهای مولکولی چندزیرواحدی هستند که در هر زیرواحد یک جایگاه فعال دارند و به چند سوبسترا متصل میشوند. واژه آلوستریک به معنی تغییر شکل است و کنفورماسیون این آنزیمها، با حضور سوبسترا و مولکولهای تنظیمی، بین دو حالت فعال و غیرفعال تغییر میکند.
این آنزیمها جایگاه تنظیمی دارند که به «فعالکننده» (Activator) یا «مهارکننده» (Inhibitors) متصل میشود. تغییر کنفورماسیون فعال یا غیرفعال در آنزیم، بستگی به غلظت فعالکننده و مهارکننده دارد. فعالکنندهها فعالیت آنزیم را افزایش و مهارکنندهها آن را کاهش میدهند.
بر اساس نوع سوبسترا و عامل تنظیمکننده دو نوع تنظیم آلوستریک وجود دارد که تنها مختص آنزیمها نیست.
- «تنظیم همگرا» (Homotropic Regulation): در این نوع تنظیم، سوبسترا همان فعالکننده است. مثل اتصال اکسیژن به هموگلوبین.
- «تنظیم غیرهمگرا» (Heterotropic Regulation): در این نوع تنظیم سوبسترا و عامل تنظیم متفاوت هستند. مثل اتصال کربندیاکسید به هموگلوبین.
همچنین دو مکانسیم پیشنهادی برای تنظیم آلوستریک جایگاه فعال وجود دارد.
- «مدل پیوستگی ساده» (Simple Sequential Model): در این مکانسیم فرض بر این است که تغییر کنفورماسیون در یک زیرواحد از حالت «شل» (Relax) به حالت «فشرده» (Tensed) سبب ایجاد تغییر در زیرواحدهای دیگر میشود، یعنی در یک مقطع زمانی، یک زیرواحد در حالت R و دیگری در حالت S است. این روش «همتعاملی منفی» (Negative Cooperativity) درآنزیمها را نشان میدهد.
- «مدل موزون یا متقارن» (Concerted or Symmetry Model): در این روش تنظیمی، همه زیرواحدها، همزمان در حالت R یا T قرار دارند.
نمونههایی از آنزیم آلوستریک
آنزیمهای زیادی در تنظیم مسیرهای زیستی نقش دارند که برخی از آنها بهشکل آلوستریک تنظیم میشوند. گلوکوکیناز و استیل-کوآ کربوکسیلاز از جمله این آنزیمها هستند.
- «گلوکوکیناز» (Glucokinase): این آنزیم در هومئوستازی گلوکز نقش مهمی ایفا و گلوکز را به گلوکز ۶-فسفات تبدیل میکند. تمایل این آنزیم به گلوکز کم است و در غلظت بالای گلوکز کبدی فعالیت میکند. گلوکوکیناز برای فعالیت نیاز به پروتئینها تنظیمی دارد.
- «استیل-کوآ کربوکسیلاز» (Acetyl-CoA Carboxylase): این آنزیم فرایندهای مربوط به «سنتز لیپید» (Lipogenesis) را تنظیم میکنند. فعالکننده این آنزیم، سیترات و مهارکننده آن مولکولهای اسیل-کوآ هستند. این آنزیم با فسفوریله و دفسفوریله شدنِ تحت کنترل هورمونهای گلوکاگون و اپینفرین نیز تنظیم میشود.
تغییر کنفورماسیون گیرنده برای اتصال به لیگاند
گیرندههای پروتئینی، گروهی از پروتئینهای عرض غشایی یا درونسیتوپلاسمی هستند که از بخشهای مختلفی تشکیل شدهاند و با اتصال لیگاندها از جمله هورمون، انتقالدهنده عصبی و متابولیتهای مختلف عملکرد سلول را تغییر میدهند. اتصال لیگاند به جایگاه مخصوص خود در گیرنده، تغییر کنفورماسیون گیرنده و برهمکنش بخش درونسیتوپلاسمی با مولکولهای دیگر از جمله پروتئینهای G را باعث میشود و شروع آبشاری از واکنشها را بهدنبال خواهد داشت.
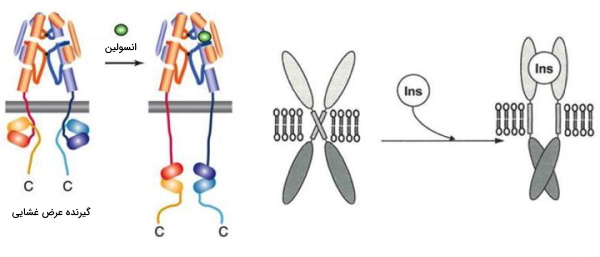
تغییر کنفورماسیون کانالهای دریچهدار
کانالهای دریچهدار به ۳ دسته تقسیم میشوند
- ولتاژی
- لیگاندی
- مکانیکی
باز و بسته شدن دریچه این کانالها به تغییرات کنفورماسیون در حضور ماده ورودی، عامل تنظیمی یا لیگاند وابسته است. برای نمونه کانالهای ولتاژی، حسگرهایی دارند که از آمینواسیدهای باردار تشکیل شده است. تغییرات پتانسیل غشا سبب حرکت این اسیدهای آمینه، تغییرات کنفورماسیون دریچه و باز شدن میشود.
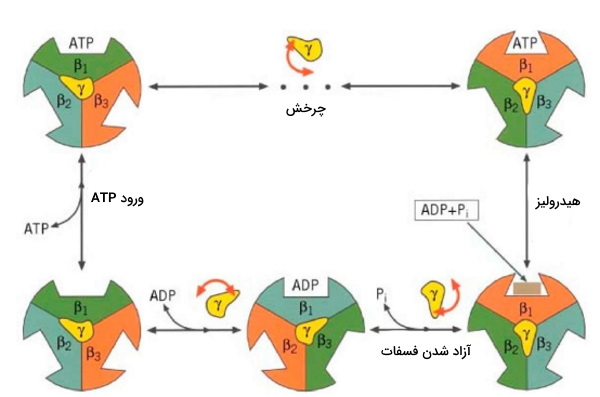
تغییر کنفورماسیون ATP سنتتاز
«ATP سنتتاز» (ATP Synthase) یا F0F1 ATPase، پمپی در غشای میتوکندری و پروکاریوتها است که با بهرهگیری از انرژی الکتروشیمیایی حاصل از انتقال یون هیدروژن، ADP را تبدیل به ATP و زنجیره فسفوریلاسیون را تمام میکند. این پمپ از دو بخش F1 (زیرواحدهای بتا، گاما و سیگما) و F0 (زیرواحدهای a، b و c) تشکیل شده است و سه کنفورماسیون عملکردی برای سنتز ATP دارد.
- «فشرده» (Tight)
- «شل» (Loose)
- «باز» (Open)
کنفورماسیون زیرواحد ، بهدنبال چرخش زیرواحد تغییر میکند. انرژی این چرخش از انتقال ، بهوسیله بخش F0 تامین میشود.
کنفورماسیونهای نوکلئیک اسید
DNA و RNA مولکولهای اطلاعاتی هستند که اطلاعات لازم برای سنتز پروتئینها را در خود ذخیره میکنند. این مولکولها بر حسب ساختار مونومری و تغییرات محیطی میتوانند ساختار مختلفی از جمله مارپیچ دوتایی، سنجاق سری و ساختارهای سه یا چهار شاخهای تشکیل دهند. در این بخش به نحوه تشکیل این ساختارها و عملکرد هرکدام در سلول میپردازیم.
ساختار دوم نوکلئوئیک اسیدها
توالی بازها در ساختار اول نوکلئوئیک اسیدها ، در تشکیل ساختار دوم نقش اساسی دارد. عامل اصلی ایجاد ساختار دوم، برهمکنش بینبازی، درون یک رشته یا بین دو رشته از DNA یا RNA است. این ساختارها شامل موارد زیر میشوند.
- مارپیچ دوتایی
- شاخهای
- حلقههای برآمده
- چهاروجهی
مارپیچ دوتایی
ویژگی ساختاری قند موجود در نوکلئوئیک اسیدها، در شکلگیری نهایی این مولکول نقش مهمی ایفا میکند. نوکلئوئیک اسیدها، چهار کنفورماسیون اصلی از مارپیچ دوتایی دارند که تفاوت آنها در قطر هلیکسی است که تشکیل شده میدهند.
- A-DNA: دو رشته موازی در جهت مخالف و مارپیچ راستگرد
- B-DNA: دو رشته موازی در جهت مخالف و مارپیچ راستگرد (مل واتسون کریک)
- Z-DNA: دو رشته موازی در جهت مخالف و مارپیچ چپگرد
- A-RNA: بخشی از RNA - دو رشته موازی در جهت مخالف و مارپیچ راستگرد
ساختارهای شاخهای
این ساختارهای موقتی در هر دو نوکلئیک اسید به دو شکل حضور دارند.
- ساختارهای «صلیبی» (Cruciforms): ساختارهای درون مولکولی
- «ساختارهای هالیدی» (Holliday Junctions): ساختارهای بین دو مولکول - دخیل در نوترکیبی
ساختارهای ۴ شاخه صلیبی در DNA، پیشنیاز آغاز همانندسازی هستند و در شکلگیری ساختار سهبعدی دیده میشوند. ساختارهای هالیدی، در فرایند تشکیل کروموزوم نوترکیب دیده میشوند. ساختارهای شاخهای، درفعالیتهای آنزیمی RNA، بسیار مهم هستند.
حلقههای برآمده
حلقههای برآمده ساختارهای DNA و RNA دورشتهای هستند. این ساختار بهدلیل وجود نوکلئوتیدهای جفتنشده در یک یا دو رشته تشکیل میشود. ممکن است ایجاد این ساختار در DNA، با اختلال در همانندسازی و رونویسی همراه باشد و به جهش مولکول منجر شود. در RNA این ساختارها در مناطقی دیده میشوند که در شناخت پروتئین و فعالیت کاتالیکی نقش دارند.
ساختارهای چهاروجهی
این ساختارها با تشکیل پیوند هیدروژنی بین چهار باز گوانین ایجاد میشوند. برخلاف مدل واتسون کریک، این پیوند بین نیتروژن ۷ از باز پورین و کربن ۶ از گروه آمینی (جفت باز هوگِستین) تشکیل میشود. این ساختارها احتمالا در انتهای غنی از گوانین تلومرها وجود دارند.
ساختار سوم نوکلئوئیک اسیدها
ساختارهای سوم در DNA و RNA، پتانسیل فعالیت آنزیمی و جایگاه اتصال به پروتئین یا کاتیونها را فراهم میکنند. یونهای فلزی با کمک نیروهای الکترواستاتیک، نقش مهمی در ایجاد ساختار سوم این مولکولها دارند. یکی از شناخته شدهترین ساختارهای سوم نوکلئیک اسید ، ساختار tRNA است. tRNA ساختاری شامل سه سنجاق سر دارد و به آن برگ شبدری میگویند. در این کنفورماسیون، بازها درگیر پیوندهای هیدروژنی درونمولکولی هستند که ساختار دوبعدی و تاخوردگیهای سهبعدی را بهوجود میآورد.
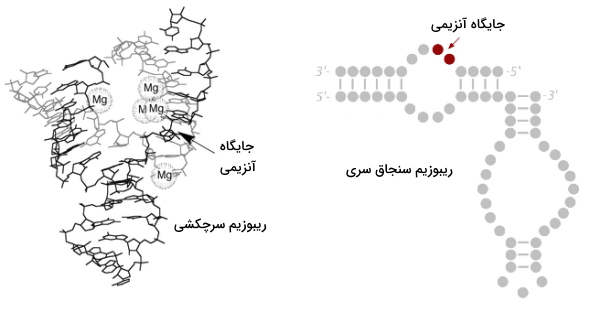
نوکلئوئیک اسیدها علاوه بر شرکت در همانندسازی، رونویسی و ترجمه، فعالیت آنزیمی نیز دارند. ریبوزیمها، RNAهایی با ساختار سهبعدی هستند که در بعضی فرایندهای بدن، نقش کاتالیزوری ایفا میکنند. دو موتیف «سرچکشی» (Hammerhead) و سنجاق سری، ساختارهای بیشتر شناخته شده در ریبوزیمها هستند. این ساختارها واکنشهای شکست و بست «نقطهمشخص» (Site Specific) برای تبدیل اولیگومر RNA حلقوی، به فرم مونومری را کاتالیز میکنند. ساختار و مکانسیم عمل این ریبوزیمها با هم متفاوت است.
- سرچکشی: از سه دومین دو رشتهای تشکیل شده است که بهوسیله بخشهای تکرشتهای بهم متصل میشوند. توالی بازی این بخشها کاملا حفظشده است.
- سنجاقسری: این ساختار از دو دومین A و B تشکیل میشود و دو طرف هر دومین ساختار هلیکس وجود دارد. تاشدگی این دومینها مجزا است اما برهمکنش ساختار سهبعدی آنها بر عملکردی بودن کنفورماسیون این ریبوزیم تاثیر میگذارد.
نکته: موتیف گروهی از ساختارهای دوم یک ماکرومولکول و دومین واحد عملکردی آن است.
DNA فراپیچش
کنفورماسیون «فراپیچش» (Supercoiled) نشان دهنده پیچخوردگیهای فراوان مولکول دو رشته DNA است که به فشردگی مولکول و قرارگرفتن آن در سلولهای پروکاریوتی و یوکاریوتی کمک میکند. B-DNA در حالت «شل» (Relax) حول محور عمودی، پیچخوردگیهایی به فاصله ۱۰٫۴ تا ۱۰٫۵ جفتباز دارد که هر هلیکس را تشکیل میدهد. ساختار فراپیچش، با ایجاد پیچخوردگیهای بیشتر در مولکول، تعداد جفتبازهای موجود در هر هلیکس را افزایش میدهد. در سلولهای پروکاریوتی این کنفورماسیون بهکمک آنزیم «ژیراز» (Gyrase) و در یوکاریوتها به کمک آنزیم «توپوایزومراز» (Topoisomerase) ایجاد میشود.
آنالیز کنفورماسیون مولکولهای زیستی
بیشترین عملکرد موجودات زنده به کمک پروتئینها انجام میگیرد. برای اینکه عملکرد کامل یک پروتئین مثلا فسفوریلاز در فسفوریلاسیون را متوجه شویم، به اطلاعات در مورد ساختار بیوشیمیایی این مولکول نیاز داریم. در این بخش روشهایی برای تعیین کنفورماسیون پروتئینها و نوکلئوئیک اسیدها را شرح میدهیم.
- «پراش پرتو ایکس» (Diffraction of X-ray)
- رزونانس مغناطیسی هسته (Nuclear Magnetic Resonance|NMR)
پراش پرتو X
پیشبینی ساختار دوم پروتئین بهکمک توالی آمینواسیدی و ساختار نوکلئوئیک اسیدها بهکمک بازهای آلی آن امکان پذیر است. اما امکان پیشبینی ساختار سوم پروتئین تنها در صورتی فراهم میشود که عملکرد آن بسیار شبیه مولکولی باشد که ساختار سهبعدی آن از قبل مشخص شده است. یکی از معمولترین روشهایی که برای تعیین ساختار سهبعدی مولکولها بهکار میگیریم، روش پراش پرتو X است. در این روش مراحل زیر انجام میشود.
- پرتو باریک X را به مولکول پروتئین یا نوکلئیک اسید خالص میتابانیم.
- بیشتر پرتوها از مولکول عبور میکنند و بخش کمی از آنها، بهوسیله اتمهای موجود در مولکول پراکنده میشوند.
- اگر نمونه مورد مطالعه، کامل کریستاله شده باشد، پرتوهای پراکنده شده در یک نقطه یکدیگر را تقویت میکنند.
- نقاط تقویتشده توسط آشکارساز ضبط و نشان داده میشوند.
آنالیز الگوی پراش، نقشه پیچیدهای از چگالی الکترونها فراهم میکند. تفسیر این نقشه، کاری بسیار پیچیده است و برای انجام آن، باید توالی آمینواسیدی پروتئین و بازهای نوکلئوئیک اسیدها را بدانیم. اخیرا دانشمندان، از این روش برای تعیین ساختار مولکولهای بسیار بزرگ ریبوزوم با بیش از ۵۰ پروتئین و چند RNA، نیز بهره بردهاند.
رزونانس مغناطیسی هسته
مولکولهای هیدروژن خاصیت مغناطیسی لحظهای دارند که از مدتها قبل، در روش NMR برای تعیین ساختار مولکولهای مختلف استفاده میشود. امروزه این روش به تعیین ساختار و کنفورماسیون پروتئینها و نوکلئوئیک اسیدها نیز کمک میکند. NMR برای مولکولهایی با وزن و کمتر مناسب است و برای مولکولهای بزرگتر وضوح مناسبی ندارد. برای انجام این روش به محلول غلیظ نمونه نیاز داریم. محلول بودن نمونه، شرایط دنبال کردن تغییرات کنفورماسیونی پروتئین در فرایندهایی مثل تاخوردگی و اتصال به پیشماده را فراهم میکند.
روش داکینگ و پیشبینی تغییر کنفورماسیون پروتئین
«داکینگ مولکولی» (Molecular Docking) روشی محاسباتی برای پیشبینی ساختار لیگاند-رسپتور و تغییرات کنفورماسیونی این دو مولکول پس از اتصال است. این روش امکان بررسی رفتار داروها در جایگاه اتصال مولکول هدف را براساس اصول بیوشیمیایی فراهم میکند. داکینگ مولکولی در حال حاضر با فرض انعطاف لیگاند و عدم انعطاف رسپتور انجام میشود و از دو قدم اولیه تشکیل شده است.
- پیشبینی کنفورماسیون لیگاند و تغییر آن در جایگاه اتصال
- ارزیابی تمایل اتصال بین لیگاند و رسپتور

جمعبندی
مولکولهای زیستی از سطوح مختلف ساختاری تشکیل شدهاند که شناخت آنها اساس زیست شناسیمولکولی را تشکیل میدهد. ساختار اولیه این مولکولها شامل نوع و توالی زیرواحدها میشود که در ساختارهای بعدی و جهتگیری فضایی مولکول بسیار موثر است. کنفورماسیونهای مختلف یک مولکول زیستی مربوط به ساختارهای سهبعدی آن است که در برهمکنش با دیگر مولکولها تغییر میکند و عملکرد مولکول را نیز تغییر میدهد.









