



انتقال فعال سلولی چیست؟ – به زبان ساده + فرق انتقال فعال اولیه و ثانویه
سلول کوچکترین واحد سازنده همه موجودات است و برای ادامه بقای خود، پایداری ساختار و انجام فرایندهای زیستی نیاز به دریافت مواد از محیط و خارج کردن مواد زائد از سیتوپلاسم دارد. برای رسیدن به یا خارج شدن از سیتوپلاسم، مولکولها باید از سد غشای پلاسمایی عبور کنند. بعضی مولکولهای کوچک به راحتی و بدون نیاز به کمک، از بین فسفولیپیدهای غشا عبور میکنند. اما مولکولهای بزرگتر برای عبور از غشا نیاز به کمک دارند. انتقال فعال یکی از مکانیسمهایی است که در سلولهای پروکاریوتی و یوکاریوتی برای انتقال مواد استفاده میشود. در این مطلب انواع انتقال فعال و مکانیسمهای مختلف آن را توضیح میدهیم.
- خواهید آموخت غشای سلولی و پمپها چه نقشی در جابهجایی انتخابی مواد دارند.
- یاد میگیرید تفاوت انتقال فعال اولیه و ثانویه را توضیح دهید.
- میآموزید شیب غلظت و شیب الکتریکی چگونه حرکت یونها را تعیین میکنند.
- نحوه عملکرد پمپ سدیم-پتاسیم و پیامدهای آن روی سلول را یاد میگیرید.
- با ساختار و کاربرد پمپهای کلاس «P» ،«V» ،«F» و «ABC» آشنا میشوید.
- خواهید آموخت چگونه نقص پروتئینهای انتقال میتواند منجر به اختلالات سلولی شود.


انتقال فعال چیست ؟
غشای سلولی از مولکولهای دوگانه دوست فسفولیپید تشکیل شده است. این مولکولها یک سر قطبی دارند که سمت سیتوپلاسم سلول و ماتریکس خارج سلولی قرار میگیرد و اسیدهای چرب آبگریز بخش میانی را میسازند. این ساختار غشا سبب میشود سدی بین فضای بیرون و خارج از سلول ایجاد میکند که به همه مواد اجازه عبور نمیدهد. مولکولهای کوچک و محلول در چربی در جهت شیب غلظت و بهوسیله انتشار ساده از غشای پلاسمایی عبور میکنند. اما انتقال مولکولهای بزرگ و قطبی نیاز به ناقلهای پروتئینی و مصرف انرژی دارد. انتقال فعال، یکی از مکانیسمهای سلولها برای وارد و خارج کردن مواد از سیتوپلاسم است. این مکانیسم مواد را برخلاف شیب غلظت یا شیب الکتریکی بین سیتوپلاسم و ماتریکس خارج سلولی منتقل میکند.
پیشنهاد میکنیم برای یادگیری انتقال فعال و غیرفعال فیلم آموزش رایگان انتقال فعال و غیرفعال در غشای سلولی در زیست دهم در فرادرس را تماشا کنید. برای تماشای این فیلم آموزشی، کافی است ویدئوی زیر را پخش کنید.
شیب غلظت
شیب غلظت دو طرف غشا، زمانی ایجاد میشود که غلظت یک ماده در سیتوپلاسم یا ماتریکس خارج سلولی بیشتر است. این اختلاف غلظت سبب حرکت مولکول از غلظت بالا به غلظت پایین میشود و این انتقال تا زمانی که غلظت مولکول در دو طرف به تعادل برسد ادامه خواهد داشت. انتشار ساده و انتشار تسهیل شده دو مکانیسم انتقالی از غشای سیتوپلاسمی هستند که بر اساس شیب غلظت مواد را منتقل میکنند. مواد محلول در چربی (هورمونهای استروئیدی) یا مولکولهای بسیار کوچک (مولکولهای گاز) بهوسیله انتشار ساده از بین مولکولهای فسفولیپید عبور میکنند و وارد سلول میشوند. اما مواد بزرگتر و قطبی ازجمله (گلوکز) بهوسیله انتشار تسهیل شده و با مشارکت ناقلهای غشایی منتقل میشوند.
شیب الکتریکی
تعداد زیادی یون در سیتوپلاسم و ماتریکس خارج سلولی وجود دارد که نوع و مقدار بار الکتریکی آنها با هم متفاوت است. این اختلاف بار مثل اختلاف غلظت، شیب الکتریکی در دو طرف غشای پلاسمایی ایجاد میکند که در انتقال برخی مواد نقش دارند. پتانسیل الکتریکی (ولتاژ) غشاهای زیستی، نشاندهنده این اختلاف الکتریکی است. در سلولهای جانوری سیتوپلاسم سلول نسبت به ماتریکس خارج سلولی بار مثبت کمتری دارد و ولتاژ آن نسبت به خارج از سلول کمتر است. پتانسیل الکتریکی غشای سلولهای جانوری بین ۴۰- و ۸۰- میلیولت تغییر میکند. یون پتاسیم و سدیم دو عامل مهم برای تنظیم پتانسیل الکتریکی غشاهای زیستی هستند. در حالت طبیعی غلظت یون پتاسیم () درون سلول بیشتر از غلظت یون سدیم () است.
- غلظت یون سدیم درون سیتوپلاسم کمتر از ماتریکس خارج سلولی و پتانسیل الکتریکی آن منفیتر است. در نتیجه شرایط به نفع ورود یون سدیم به سیتوپلاسم عمل میکند.
- شیب الکتریکی یون پتاسیم خارج سلولی در جهت ورود این یون به سیتوپلاسم اما غلظت بیشتر داخل سلول در جهت خروج آن از سلول عمل میکند. غلظت نهایی پتاسیم در دو طرف غشا به تعادل این دو نیرو بستگی دارد.
معادله نرست برای غشای زیستی
معادله نرست فرمولی برای محاسبه پتانسیل لازم برای انتقال یک یون خلاف جهت شیب الکتریکی یا غلظت است. در بخشهای قبلی توضیح دادیم که کاتیونهای و آنیونهای دو طرف غشای زیستی تمایل دارند از بخش با غلظت (بار الکتریکی) بیشتر به بخش با غلظت کمتر حرکت کنند. سرعت این انتقال در ابتدا بیشتر است و رفتهرفته کاهش مییابد. با به تعادل رسیدن غلظت (بار الکتریکی) یونها در دو طرف غشا، سرعت انتقال صفر خواهد شد.
شرایطی را در نظر بگیرید که غظت یک کاتیون مشخص داخل سلولی بیشتر از خارج سلول، اما بار مثبت کلی خارج از سلول بیشتر از داخل سلول است. در این وضعیت یونها به کدام جهت منتقل میشوند؟ پتانسیل نرست فرمولی است که با اندازهگیری پتانسیل تعادل، جهت انتقال یونها را از فرمول زیر تعیین میکند.
در این معادله R ثابت گازها، T دما، F ثابت فارادی، z بار یون، غلظت یون خارج از سلول و غلظت یون داخل سلول است. اگر R را ۸٫۳۱، دما را ۳۷ درجه سانتیگراد (۳۱۰٫۲ کلوین) در نظر بگیریم، ln را به log () و ولت را به میلیولت (واحد غشاهای زیستی) تبدیل کنیم، فرمول نرست به شکل زیر تغییر میکند.
تصور کنیم غلظت سلول ۱۰۰ و غلظت خارج سلولی یون ۵ میلی مولار باشد. پتانسیل تعادل یون را میتوان از از معادله زیر محاسبه کرد.
در نتیجه اختلاف پتانسیل تعادل برای یون سدیم در دو طرف غشا، باید ۸۰- میلیولت باشد. اگر اختلاف پتانسیل ۵۰- میلیولت باشد، در این شرایط یون سدیم در جهت شیب غلظت از سلول خارج میشود. اما اگر اختلاف پتانسیل دو طرف غشا ۱۰۰- باشد، یونها بر خلاف شیب غظت وارد سلول میشوند.
انواع انتقال فعال
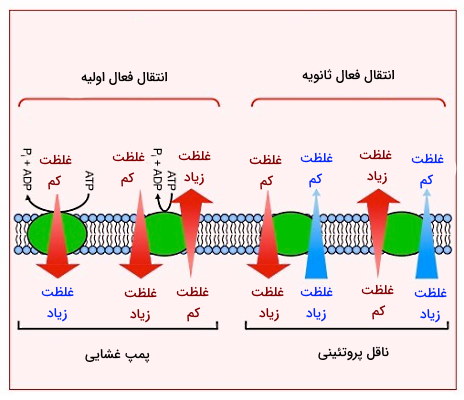
برای انتقال مواد در جهت خلاف شیب غلظت و شیب الکتریکی، نیاز به مصرف انرژی وجود دارد. به همین دلیل بخش زیادی از ATP سلولها در انتقال فعال مواد از عرض غشا مصرف میشود. انتقال فعال اولیه و همانتقالی یا انتقال فعال ثانویه انواع انتقال فعال در غشاهای زیستی هستند که در این بخش به توضیح آن میپردازیم.
انتقال فعال اولیه
انتقال فعال اولیه به کمک پروتئینهای عرض غشایی به نام پمپ انجام میشود. در این نوع انتقال ATP، نور خورشید (فتوسنتز گیاه یا باکتری) یا واکنشهای اکسایس-کاهش انرژی لازم برای انتقال مواد خلاف جهت شیب غلظت یا الکتریکی را فراهم میکند. پمپها از زنجیرههای پلیپپتیدی تشکیل میشوند که هر کدام چند بار از غشا عبور میکنند. در این بخش ساختار و عملکرد 5 نوع پمپ در سلولهای گیاهی، جانوری و باکتری را بررسی میکنیم.
- پمپ وابسته به نور
- پمپهای ATPase
- پمپ کلاس V
- پمپ کلاس F
- پمپ ABC
پمپ نوری باکتریورودوپسین
باکتریوروپوپسین، پمپ پروتونی با چند زنجیره پلیپپتیدی در غشای آرکیباکترها هالوفیل (نمکدوست) است و از انرژی نور خورشید برای انتقال پروتون استفاده میکند. زنجیرههای پپتیدی این پمپ از ۷ آلفا هلیکس عرض غشایی تشکیل میشود. رتینال (آلدهید ویتامین A) بخش جذبکننده نور (کروموفور) در این پمپ است که با پیوند کووالانسی به زنجیره جانبی باقیمانده لیزین متصل شده است. پروتون برای خروج از سلول به ترتیب به باقیمانده آسپارتات ۹۶، رتینال و آسپارتات ۸۵ در سمت سیتوپلاسمی پمپ متصل میشود. جذب نور بهوسیله رتینال، کنفورماسیون پروتئینها را تغییر میدهد و پروتون منتقل میشود.
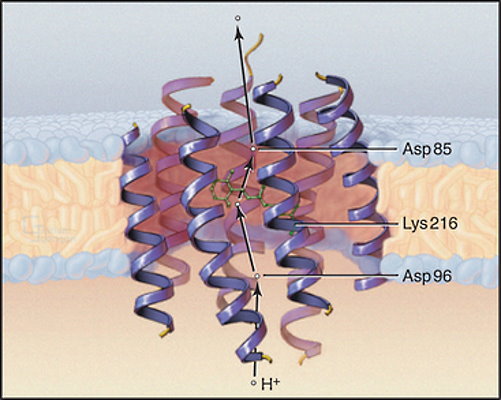
در مرحله بعد ATP سنتتاز پروتونی، از اختلاف پتانسیل الکتریکی ایجاد شده بهوسلیه باکتریورودوپسین برای تولید انرژی شیمیایی موجود در ATP استفاده میکند.
پمپ ATPase
سه کلاس مختلف از ATPase های غشایی تبادل مواد، خلاف جهت شیب غلظت را تسهیل میکنند که ساختار و عملکرد آنها با هم متفاوت است. اما تمام آنها پروتئینهای عرض غشایی هستند که از بخشهای هیدروفوب و هیدروفیل تشکیل میشوند.
پمپهای ATPases نوع P
پمپهای ATPase نوع P یا گروهی از ناقلهای پروتئینی هستند که در باکتریها، آرکیباکتریها و یوکاریوتها به انتقال یونها و مولکولهای لیپیدی از عرض غشا کمک میکنند. ساختار این پمپها از آلفا هلیکسهای عبوری از غشا تشکیل شده است و باقیمانده آسپارتات در جایگاه فعال کلاسهای مختلف این ناقلهای آنزیمی وجود دارد. به علاوه در تمام این پمپها تغییر کنفورماسیون بین دو حالت و در هیدرولیز ATP و انتقال یونها نقش دارد.
ساختار تمام پمپهای p یکسان است و از دومینهای M (عرض غشایی)، P (جایگاه فسفوریلاسیون)، N (جایگاه اتصال نوکلئوتید)، A (فعالکننده) و R (تنظیمی) تشکیل میشود.
- دومین عرض غشایی: این بخش از پمپ معمولا شامل ۱۰ تا ۱۲ آلفا هلیکس عرض غشایی است که جایگاه اتصال به لیگاندها در نزدیکی بخش میانی آن قرار دارد. ۶ دومین عرض غشایی در انتقال لیگاند نقش دارند و ۴ تا ۶ دومین دیگر به حفظ ساختار پمپ کمک میکنند.
- دومین فسفوریلاسیون: باقیمانده آسپارتیک اسید در این بخش قرار دارد که جایگاه آن در تمام انواع این پمپ یکسان است. دومین P به دو بخش تقسیم میشود که توالی آمینواسیدی آن بسیار متفاوت است. این دو بخش، یک ساختار بتاشیت موازی با ۷ رشته که ۸ آلفا هلیکس اظراف آن قرار دارد.
- دومین اتصال نوکلئوتید: دومین N یک کیناز ساختاری است که بین دو بخش دومین P قرار و از ساختار بتا شیت غیرموازی با ۷ رشته و دو زنجیره آلفا هلیکس تشکیل شده است.
- دومین فعالکننده: دومین A کوچکترین دومین سیتوپلاسمی در این پمپها است که فعالیت فسفاتازی دارد و با هیدرولیز فشفات اضافه شده به دومین P کنفورماسیون پمپ را تغییر میدهد.
- دومین تنظیمی: دومین R در بعضی از پمپهای کلاس P وجود دارد. برای مثال انتهای C در پمپهای کلسیم غشای سلولهای ماهیچهای جایگاه تنظیمی برای پروتئین کلمودولین دارد که در حضور یون کلسیم پمپ را فعال میکند.
پمپهای کلاس P بر اساس توالی آمینواسیدهای ساختار به ۵ گروه اصلی تقسیم میشوند. P1A ATPases پمپهایی هستند که یون پتاسیم را به سیتوپلاسم سلولها منتقل میکنند. P1B ATPases پمپهای انتقال فلزات سنگین ازجمله روی، مس و نقره در غشای پروکاریوتها هستند. P2A ATPases ( و ،) ناقلهای عرض غشایی سلولهای یوکاریوتی هستند که در انتقال یونهای سدیم، پتاسیم، پروتون، کلسیم، منیزیم و فسفولیپیدها شرکت میکنند. P3 ATPases به دو گروه اصلی A و B تقسیم میشوند. گروه A پمپهای پروتون () در غشای سلولهای یوکاریوتی، گیاهی و قارچها هستند و گروه B در انتقال منیزیم از غشای یوباکتریها و گیاهان شرکت میکنند.
انواع مختلف این پمپ در بافتهای انسان در ایجاد پتانسیل عمل نورونهای عصبی، استراحت ماهیچهها، ترشح و بازجذب در نفرونهای کلیه و جذب مواد در روده کوچک شرکت میکنند. پمپ سدیم-پتاسیم، پمپ پروتون-پتاسیم، پمپ کلسیم و پمپ پروتون گیاهان و قارچها انواع مختلف P- ATPase هستند که عملکرد آنها را در این بخش بررسی میکنیم.
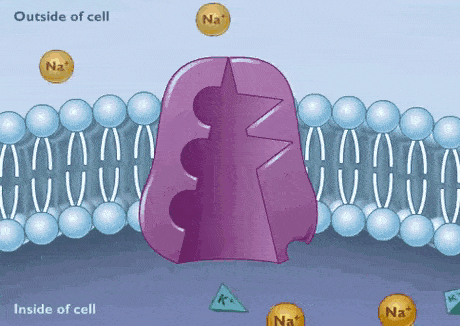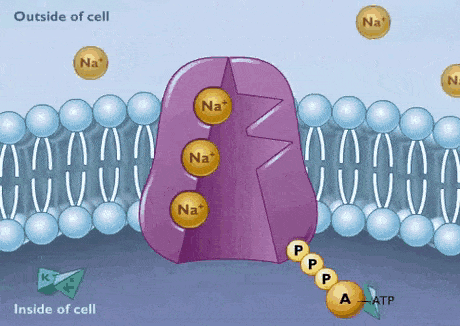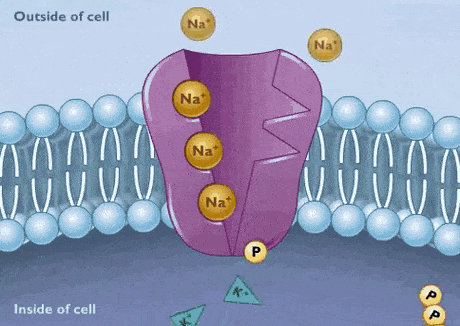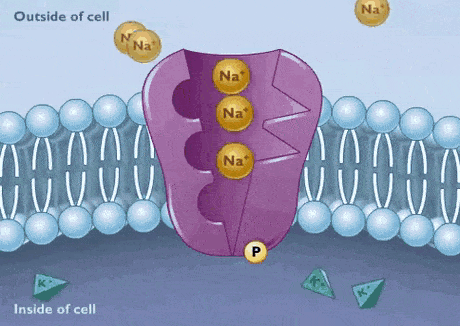
پمپ سدیم پتاسیم
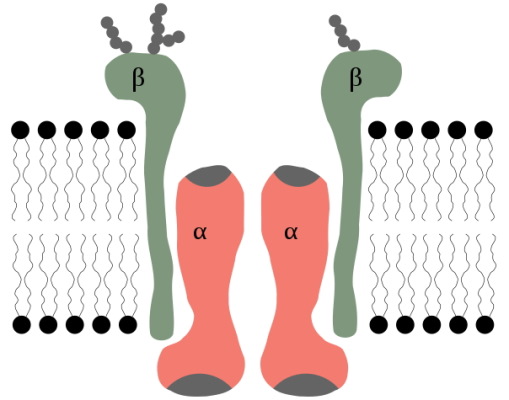
پمپ سدیم-پتاسیم یکی از ATPaseهای نوع P است که در غشای تمام سلولهای جانوری وجود دارد و با هیدرولیز هر مولکول ATP، سه یون سدیم را از سلول خارج و دو یون پتاسیم را وارد سلول میکند. در نتیجه به ازای هر ATP یک بار الکتریکی مثبت از سلول خارج میشود. ساختار این پمپ از دو زیر واحد آلفا و دو زیرواحد بتا تشکیل شده است.
- زیر واحد آلفا: از ۱۰ آلفا هلیکس عرض غشایی تشکیل شده است. که سه جایگاه اتصال به دارد. این زیرواحد در کنفورماسیون به یونها متصل میشود.
- زیر واحد بتا: دو جایگاه اتصال به در این زیرواحد قرار دارد و در کنفورماسیون به یونها متصل میشود.
این پمپ در سلولهای مختلف با انتقال یونها، پتانسیل استراحت غشای پلاسمایی و حجم سلول را تنظیم میکند. مهار منجر به تجمع یون سدیم داخل سیتوپلاسم، افزایش فشار اسمزی داخل سلول، ورود آب به سلول، افزایش حجم آن و در نهایت پلاسمولیز سلول میشود. به علاوه این پمپ در مسیرهای پیامرسانی MAPK، سنتز رادیکالها آزاد میتوکندری و تنظیم کلسیم داخل سیتوپلاسم (سیگنالینگ IP3 و فسفولیپاز C) شرکت میکند. این پمپ در فعالیتهای زیستی مختلفی شرکت میکند.
- میلیونها پمپ سدیم-پتاسیم در غشای هر سلول اپیتلیال در لوله پیچخورده دور نفرون وجود دارد که برای بازجذب آمینواسیدها، بازجذب گلوکز، تنظیم الکترولیت و pH خون ضروری است.
- اختلاف پتانسیل ایجاد شده بهوسیله پمپ سدیم-پتاسیم برای حرکت اسپرم در لولههای خروجی بیضه و فعالیت آکروزوم در فرایند الحاق با تخمک ضروری است.
- فعالیت این پمپ در نورونها، غشا را از حالت پلاریزه به حالت استراحت برمیگرداند. به علاوه فعالیت این پمپ در غشای آستروسیت به حفظ شیب الکتروشیمیایی یون سدیم و بازجذب نوروترانسمیترها از شکاف سیناپسی کمک میکند.
- خارج شدن سدیم از سلول انرژی لازم برای انتقال فعال ثانویه سایر مولکولها ازجمله گلوکز و آمینواسید را تامین میکند.
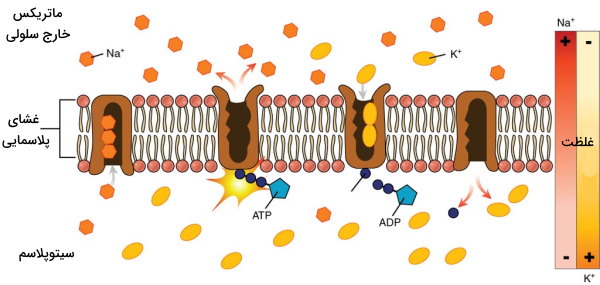
تمایل زنجیره پلیپپتیدی پمپ به بیشتر از است. به همین دلیل ابتدا سه یون سدیم در سمت سیتوپلاسمی به زیرواحد آلفای پمپ متصل میشوند. در مرحله بعد، ATP هیدرولیز و باقیمانده آسپارتات را فسفوریله میکند. فسفوریله شدن پمپ با تغییر کنفورماسیون سبب کاهش تمایل زنجیره پلیپپتیدی به و در آشکار شدن جایگاه اتصال به میشود. در نتیجه یونهای سدیم از زیرواحد آلفا جدا و وارد ماتریکس خارج سلولی میشود و یون پتاسیم به زیرواحد بتا متصل خواهد شد. این اتصال با دفسفوریله شدن پمپ، تغییر کنفورماسیون به حالت و ورود به سیتوپلاسم همراه است.
پمپ کلسیم
یکی دیگر از پمپهای کلاس P، پمپهای کلسیم هستند که با انتقال فعال از سیتوپلاسم بسیاری از فرایندهای سلولی ازجمله سیگنالینگ و انقباض ماهیچهها را کنترل میکنند. درصد زیادی از پروتئینهای غشای شبکه اندوپلاسمی صاف در سلولهای ماهیچهای را این پمپ تشکیل میدهد که با هیدرولیز ATP غلظت یونهای کلسیم سارکوپلاسم را کاهش میدهد. در نتیجه میوزین غیرفعال و ماهیچه و انقباض ماهیچه مهار میشود. این ناقل پروتئینی از سه دومین سیتوپلاسمی و یک بخش هیدروفوب عرض غشایی با ۱۰ هلیکس آلفا تشکیل شده است. دو جایگاه اتصال یون کلسیم در بخش عرض غشایی قرار دارد و با مکانیسم زیر ر از سلول خارج میکند.
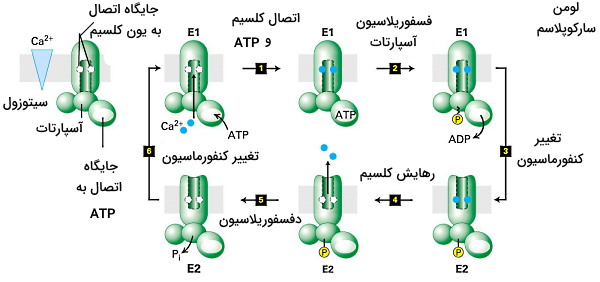
کنفورماسیون تمایل بیشتری برای اتصال به و ATP دارد. در نتیجه هیدرولیز ATP، فسفوریله شدن باقیمانده آسپارتات و تغییر کنفورماسیون ۲ یون کلسیم از سلول خارج و ۲ پروتون وارد میشوند. دفسفوریله شدن زنجیره پلیپپتیدی، کنفورماسیون پمپ را به وضعیت برمیگرداند و چرخه انتقال کلسیم ادامه دارد.
پمپ هیدروژن پتاسیم
تنظیم pH معده یکی از مهمترین فعالیتهایی است که بهوسیله پمپ هیدروژن-پتاسیم در سلولهای دیواره معده انجام میشود. این پمپ نوع P در غشای سلولهای جانبی مخاط معده با انتقال هر به سیتوپلاسم یک هیدرونیوم () را به لومن معده منتقل میکند. به همین دلیل برخلاف سایر ATPaseها بار الکتریکی منتقل نمیکند.
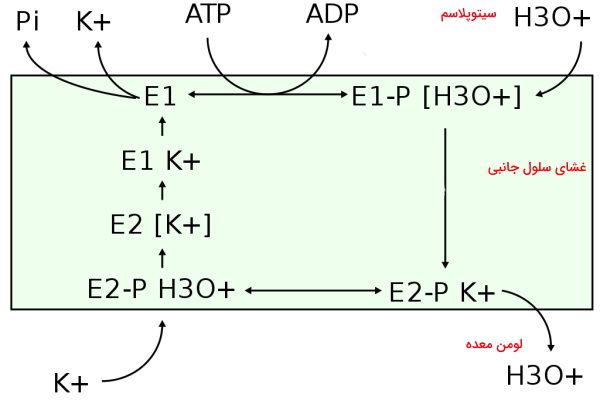
ترشح گاسترین از سلولهای G معده، ترشح هورمون هیستامین از سلولهای شبه انتروکرومافین را تحریک میکند. اتصال هیستامین به گیرندههای در غشای سلولهای جانبی با فعال کردن مسیر cAMP سبب انتقال وزیکولهای ترشحی به غشا و ورود قرار گرفتن پمپ هیدروژن-پتاسیم در غشا میشود.
انتقال فعال لیپیدها
انتقال لیپیدها بین دو لایه غشای لیپیدی بهوسیله سه گروه از پروتئینهای عرض غشایی انجام میشود. P4-ATPases یا فلیپاز تنها پمپهای کلاس P هستند که در انتقال لیپیدهای غشایی نقش دارند که ژن کدکننده آن تنها در یوکاریوتها وجود دارد. این ناقلها با هیدرولیز ATP، فسفولیپدها را بین دو لایه غشا جابهجا میکنند. عملکرد این پمپ سبب ایجاد و حفظ عدم تقارن در غشای پلاسمایی میشود و برای انجام بسیاری از فرایندهای سلولی ازجمله جوانه زدن وزیکولها، سیگنالینگ سلول، انعقاد خون، هومئوستازی کلسترول و صفرا، آپوپتوز و زنده ماندن نورونها ضروری است. بعضی از P4-ATPases مولکولهای فسفاتیدیل سرین و فساتیدیل اتانول آمین را بین دو لایه غشا جابهجا میکنند. اما گروهی از این پمپها به انتقال فسفاتیدیل کولین اختصاص یافتهاند. فسفوریله شدن باقیمانده آسپارتات در ساختار این پمپ مثل سایر پمپهای کلاس P سبب تغییر کنفورماسیون زنجیرههای پلیپپتیدی و انتقال لیپید از لایه خارجی به لایه سیتوپلاسمی غشا میشود.
این پمپها به پنج گروه تقسیم میشوند که ساختار همه آنها از زنجیره پلیپپتیدی بسیار بزرگی (۱۲۰ kDa) با چهار دومین مختلف تشکیل شده است. بر اساس توالی آمینواسید سه ناحیه سیتوپلاسمی در این زنجیره وجود دارد که در فعالیت آنزیمی پمپ شرکت میکنند. تفاوت این پمپ با سایر پمپهای کلاس p وجود مجموعه پروتئین CDC50 کنار ساختار اصلی است که به اتصال لیپید و پمپ کمک میکند. بخشهای عرض غشایی M1-M6 اجرای اصلی بخش انتقالی و M7-M10 بخشهای کمکی هستند.
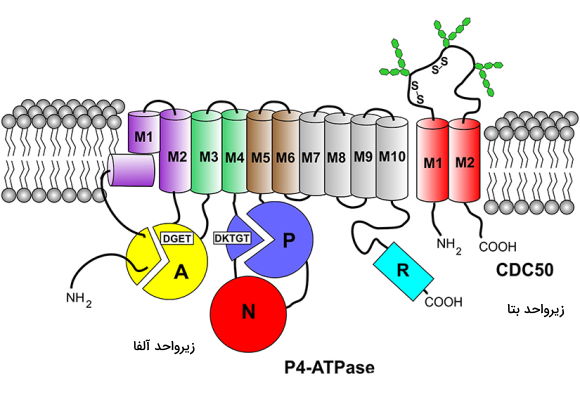
«فلوپازها» (Floppase) گروه دیگری از ناقلهای فسفولیپید غشای پلاسمایی هستند که لیپیدها را از لایه داخلی به لایه خارجی انتقال میدهند. این ناقلها از خانواده پمپهای ABC هستند که با مصرف دو مصرف ATP برای انتقال مواد استفاده میکنند. MsbA یکی از انواع این ناقل در غشای باکتری گرم منفی ای. کلی است که لیپید A را به سطح خارجی غشا انتقال میدهد.
پمپ ATPase نوع V
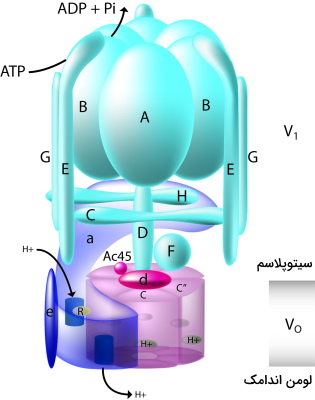
پمپ ATPase نوع V یا پمپ واکوئلی این ناقل عرض غشایی در غشای پلاسمایی اندامکها (اندوزومها، لیزوزوم و وزیکولهای ترشحی) و غشای سلولی بعضی سلولهای یوکاریوتی وجود دارد. این پمپ با شکستن پیوندهای ATP در دو طرف غشای زیستی اختلاف غلظت یون هیدروژن ایجاد میکند. این پمپ در غشای وزیکولهای سیناپسی با افزایش غلظت یون هیدروژن، pH داخلی وزیکول را کاهش میدهد. این پمپ از دو زیرواحد اصلی تشکیل شده است که هر کدام شامل بخشهای مختلفی میشوند.
- زیرواحد V1: جایگاه هیدرولیز ATP در این زیرواحد هیدروفیل قرار دارد. این پمپ از یک هگزامر با زنجیرههای پلیپپتیدی A و B، یک بخش مرکزی D و زیرواحدهای D و E محیطی و بخشهای تنظیمی C و H تشکیل شده است. هیدرولیز ATP در زیرواحدهای AB تغییر کنفورماسیون ایجاد میکند و سبب چرخش زیرواحد D میشود.
- زیرواحد V0: ناقل پروتون در این زیرواحد قرار دارد و برخلاف پمپهای کلاس F پروتون را خلاف جهت شیب غلظت الکتروشیمیایی از عرض غشا عبور میدهد. زنجیرههای پلیپپتیدی تشکیل دهنده این زیرواحد، هیدروفوب هستند.
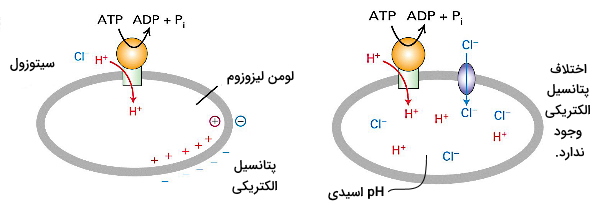
بسیاری از پمپهای این کلاس برای حفظ تعادل الکتروشیمیایی دو طرف غشا با پمپهای میشوند. تصویر شماتیک زیر عملکرد پمپ V در غشای لیزوزوم را نشان میدهد. همزمان با ورود بهوسیله پمپ، بهوسیله کانالها وارد لیزوزوم میشود. اگر همراهی یون کلر نباشد، پس از مدتی اختلاف پتانسیل الکتری در دو طرف غشا مانع از ورود بیشتر به لیزوزوم میشود و فعالیت این اندامک را مختل میکند.
F0F1-ATPases
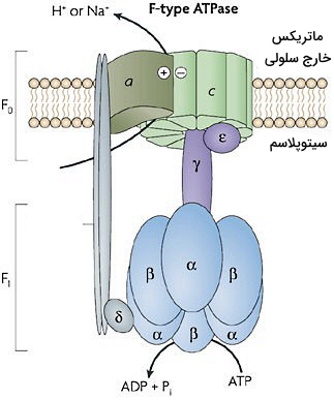
پمپ F0F1-ATPases یا F0F1 آز-سنتتاز ناقل پروتئینی در غشای باکتریها، غشای داخلی میتوکندری (کمپلکس V) و غشای تیلاکوئیدی کلروپلاست است. این پمپ با انتقال یون هیدروژن در جهت شیب غلظت الکتروشیمیایی و با اضافه کردن فسفات، ADP را به ATP تبدیل میکند. این پمپ از دو زیرواحد اصلی تشکیل شده است.
- زیرواحد F۱: این زیرواحد هیدروفوب در غشای داخلی میتوکندری قرار دارد و از سه زیرواحد و سه زیر واحد تشکیل میشود. چرخش زیرواحد این بخش با تغییر کنفورماسیون زنجیرههای پلیپپتیدی و تولید ATP همراه است. در هر چرخش کامل پمپ، سه کنفورماسیون متفاوت در زیرواحد بتا ایجاد میشود که شرایط فضایی لازم برای اتصال ADP به جایگاه فعال و خروج ATP از این جایگاه را فراهم میکند. باقیمانده آمینواسیدی Gly161 ، Tyr345، Phe418 و مولکول آب با ایجاد پیوند هیدروژنی باز آدنین را در جایگاه فعال نگه میدارند. ریبوز با Ser372 و Arg373 پیوند هیدروژنی برقرار میکند. چرخش ۱۲۰ درجه زیر واحد گاما موقعیت فضایی برای اتصال ADP و فسفات معدنی را فراهم میکند. چرخش و تغییر کنفورماسیون بعدی، پیوندهای هیدروژنی بین جایگاه فعال و ATP تشکیل شده را میشکند و این نوکلئوتید از جایگاه فعال خارج خواهد شد.
- زیرواحد F۰: این زیرواحد هیدروفیل در ماتریکس میتوکندری قرار دارد. زیرواحد B در این بخش F1 را به غشای پلاسمایی متصل میکند و بخش ثابت پمپ است. این زیرواحد از یک حلقه C (با ۹ زیرواحد داخل غشایی) و زیرواحد a تشکیل میشود. در هر یک از زیرواحدهای حلقه c یک جایگاه انتقال پروتون وجود دارد.
پمپ ABC
ناقل ABC (ATP-binding cassette transporters) گروهی از ناقلهای عرض غشایی هستند که در سادهترین سلولهای پروکاریوتی تا پیچیدهترین یوکاریوتها در انتقال مواد شرکت میکنند. این ناقلها معمولا از چند زیرواحد تشکیل میشوند که تعدادی از آنها عرض غشایی و تعدادی از آنها ATPaseهای قلاب شده به غشا هستند. بخش آنزیمی با هیدرولیز پیوند فسفودیاستری در ATP انرژی لازم برای انتقال مواد را تامین میکند. این ناقلها برخلاف پمپهایی در بخشهای قبلی توضیح دادیم، در انتقال انواع متنوعتری از مولکولهای کوچک معدنی تا مولکولهای بزرگ آلی ازجمله کربوهیدراتها، آمینواسیدها، پپتیدها، ویتامین و نوکلئوتیدها نقش دارند.
این مولکولها بر اساس جهت انتقال مواد به دو نوع اینپورتر (انتقال مواد به سیتوپلاسم) و اکسپورتر (انتقال مواد خارج از سلول) تقسیم میشوند. هر دو گروه این پمپها در غشای پروکاریوتها وجود دارد. اما تنها بسیاری یوکاریوتها تنها نوع اکسپورتر در انتقال مواد نقش دارد. ساختار اصلی این ناقلها از دو دومین NBDs (جایگاه اتصال ATP) و دو زیرواحد TMDs (ناقل لیگاند) تشکیل میشود که در یوکاریوت و پروکاریوت متفاوت هستند.
- پروکاریوت: چهار دومین ناقل ABC زیرواحدهای جدا یا یک پلیپپتید با دومینهای مختلف باشد. به علاوه ABCهای اینپورتر به یک بخش کمی نیاز دارند که در اتصال لیگاندهای محلول به زیرواحدهای TMD کمک میکند. پروتئین کمکی باکتریهای گرم منفی، پروتئین محلول در پریپلاسم و در باکتریهای گرم مثبت لیپوپروتئینی است که به لایه خارجی غشا قلاب میشود.
- یوکاریوت: بیشتر پمپهای ABC این سلولها از یک زنجیره پلیپپتیدی تشکیل شده است که چهار دومین عملکردی در آن قرا دارد و بخش های کمکی ساختار آن را حفظ میکنند.
اتصال لیگاند و ATP به جایگاه فعال زیرواحدهای NBDs منجر به تغییر کنفورماسیون این پمپ غشایی میشود. در مرحله بعد هیدرولیز ATP با تغییر دادن کنفورماسیون پمکپ سبب آزاد شدن لیگاند خواهد شد. یکی از تفاوتهای این ناقل با پمپهای دیگر در مصرف دو مولکول ATP برای انتقال هر مولکول لیگاند است. در شکل زیر بخش داخلی ناقل به لیگاند (D) متصل میشود. در مرحلع بعد، پس از اتصال دو مولکول ATP، دومینهای NBD دیمری میشوند و کنفورماسیون دومینهای TMDs را تغییر میدهند. در نتیجه لیگاند از سلول خارج میشود.
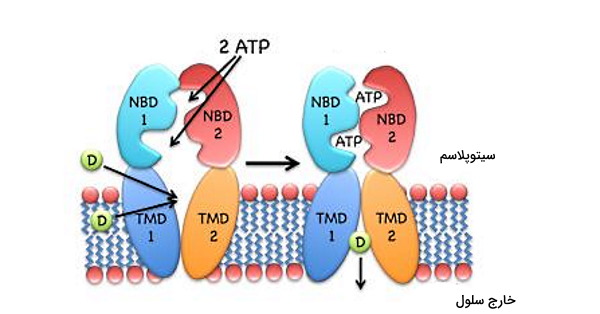
دو گروه از اینپورترهای ABC در انتقال مواد نقش دارند که تفاوت آنها نوع تغییر کنفورماسیون دومینهای TMDs است. در ناقلهای نوع I، اتصال لیگاند به بخش داخلی دومین TMDs و اتصال دو مولکول ATP و دیمری شدن NBD سبب تغییر کنفورماسیون و باز شدن کانال در جهت خارج سلول میشود. در ادامه هیدرولیز ATP موجب بازگشت کنفورماسیون به حالت اول و شروع چرخه انتقال خواهد شد.
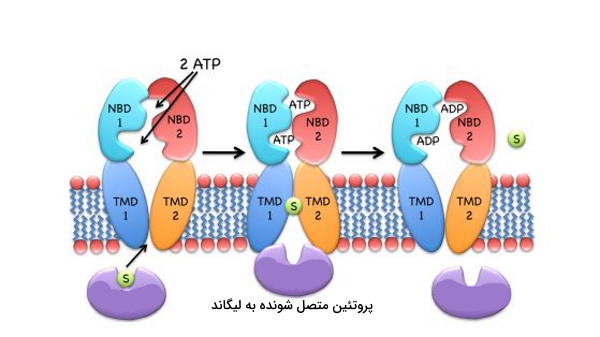
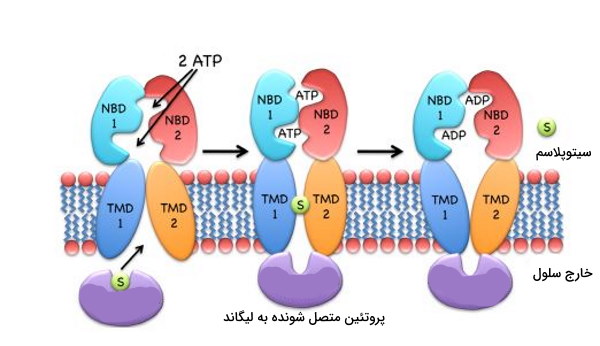
در ایمپورترهای نوع II، لیکاند پس از هیدرولیز ATP و جدا شدن دو دومین NBD از هم وارد سیتوپلاسم خواهد شد.
تفاوت پروتئین های انتقال فعال اولیه
ناقلهای انتقال فعال اولیه، همه پروتئینهای عرض غشایی هستند که از بخشهای هیدروفوب و هیدروفیل تشکیل میشوند. در پمپهای کلاس P برای تغییر کنفورماسیون و انتقال مولکولها باقیمانده آسپارتات محافظتشدهای فسفوریله میشود. سازمانیافتگی زیرواحدها در پمپهای کلاس V و F بهشکلی است که چرخش زیرواحدها حول محور عمودی سبب تغییر کنفورماسیون، هیدرولیز ATP و انتقال مواد میشود. پمپهای ABC بیشتر در غشای پروکاریوتها قرار دارند و با هیدرولیز دو مولکول ATP بسیاری از مواد معدنی و آلی را از عرض غشا عبور میدهند.
انتقال فعال ثانویه چیست ؟
شیب الکتریکی ایجاد شده بهوسیله انتقال فعال اولیه، انرژی لازم برای انجام انتقال فعال ثانویه را تامین میکند. در این انتقال یون در جهت شیب غلظت و مولکول یا یون دیگر خلاف جهت شیب غلظت از عرض غشا عبور میکند. برای مثال غلظت یون سدیم بسیار زیاد خارج از سلول را در نظر بگیرید. اگر کانال یا ناقلی در غشا باز باشد، یون سدیم به دلیل اختلاف غلظت بهراحتی و بدون مصرف انرژی وارد سلول میشود. در انتقال فعال ثانویه، انتقال سدیم بر اساس شیب غلظت با انتقال مولکول دیگری برخلاف شیب غلظت جفت میشود.انتقال «سیمپورت» (Symport) و «آنتی پورت» (Antiport) دو مکانیسم انتقال فعال ثانویه هستند. جهش در ژن این ناقلها منجر به بیماریهای مختلف ازجمله سیستیک فیبروزیس میشود.
انتقال به وسیله سیمپورتر
در این نوع انتقال یون و مولکول انتقالی در یک جهت وارد یا از سلول خارج میشوند. برای مثال گلوکز در سلولهای دیواره روده بهوسیله سیمپورترهای سدیم جذب میشود. ناقل این مولکولها، گلیوپروتئینهای عرض غشایی هستند که بخحش هیدروفوب آنها بین زنجیرههای اسید چرب دو لایه فسفولیپید قرار میگیرد. سیمپورترها به انتقال غشایی بسیاری از مواد در بافتها کمک میکنند.
- سیمپورتر -گلوکز (SGLT1) در غشای لومنی سلولهای دیواره روده کوچک و سلولهای اپیتلیال لوله پیچخورده نزدیک در نفرون، به جذب گلوکز از کیموس و بازجذب این مولکول از ادرار کمک میکند.
- سیمپورتر -فسفات یکی دیگر از پروتئینهای عرض غشایی است که از پتانسیل الکتریکی یون سدیم برای انتقال فسفات از غشای لومنی سلولهای روده کوچک و نفرونها استفاده میکند. بدون حضور این ناقل، فسفات موجود در مواد لبنی رژیم غذایی جذب نخواهد شد.
- سیمپورتر در غشای بازولترال سلولهای فولیکولی غده تیروئید، با انتقال یون ید به سیتوپلاسم سلول در تولید هورمونهای تیروئیدی نقش بسیار مهمی دارد.
- سیمپورتر (NKCC) در غشای بسیاری از بافتها به برقراری تعادل غلظت یونهای دو طرف غشای پلاسمایی به خصوص غشای غدد اگزوکرین کمک میکند. NKCC1 معمولا در غشای بازولترال (غشای نزدیک به مویرگ) قرار دارند و یک یون پتاسیم، یک یون سدیم و دو یون کلر را را همزمان از مایع میانبافتی به سیتوپلاسم منتقل میکنند. NKCC2 ناقل مهمی در بخش ضخیم و پایینرونده قوس هنله و ماکولا دنسا نفرون است و یکی از ناقلهایی است که در بازجذب سدیم شرکت میکند. هورمون ضدادراری یا وازوپرسین فعالیت این سیمپورتر را افزایش میدهد. افزایش غلظت یون سدیم در خون، اختلاف فشار اسمزی ایجاد کرده و بازجذب آب را به دنبال دارد و ادرار کاهش مییابد.
- سیمپورتر مهمترین ناقل یونی در غشای لوله پیچخورده دور نفرون است. این ناقل وظیفه بازجذب ۵ تا ۱۰ درصد مایع ترشح شده در نفرون را بر عهده دارد.
- سیمپورتر یکی دیگر از ناقلهای پروتئینی است که در بازجذب یونها از نفرون شرکت میکند. زمانی که غلظت یون پتاسیم و کلر داخل سلولی به دلیل فعالیت پمپ سدیم-پتاسیم و NKCC افزایش مییابد، این ناقل با خارج کردن پتاسیم و کلر، پتانسیل غشا را به حالت تعادل برمیگرداند.
- سیمپورتر ناقل غشایی پایانه آکسون برای انتقالدهنده عصبی GABA است که همزمان دو یون سدیم، یک یون کلر و یک مولکول GABA را به سیتوپلاسم سلول پسسیناپسی منتقل میکند.
- سیمپورتر -مونوکربوکسیلات (MCT) ناقلی است که یک مولکول لاکتات، پیرووات، استواستات یا بتا هیدروکسیبوتیرات را به همراه یک یون هیدروژن به کمک انرژی حاصل از اختلاف پتانسیل الکتریکی یون هیدروژن از عرض غشا عبور میدهد. جهت انتقال بهوسیله غلظت یون هیدروژن تنظیم میشود.
- سیمپورتر -اولیگوپپتید (PepT) یکی دیگر از ناقلهای غشای لومنی روده کوچک و یکی از سازوکارهای کمککننده به تنظیم pH لوله گوارش است. این ناقل با انتقال یک یون هیدروژن در جهت شیب غلظت به سیتوپلاسم سلولهای انتریک، یک دی یا تریپپتید حاصل از فعالیت پروتئازهای سیستم گوارش را وارد سیتوپلاسم سلولهای انتریک میکند.
- سیمپورتر سدیم-بیکربنات (NBC) یکی دیگر از ناقلهای پروتئینی است که به تنظیم اسید و باز کمک میکند. جهت انتقال یون از عرض غشا بستگی به نوع بافت و سلول دارد. برای مثال NBCe1 در غشای بازولترال لوله نزدیک در نفرونهای کلیه است و همزمان یک سه را از سلول خارج و به مایع میانبافتی منتقل میکند.
انتقال به وسیله آنتیپورتر
در این نوع انتقال، یون و مولکول انتقالی خلاف جهت هم حرکت میکنند. به دلیل عملکردی که پروتئینهای غشایی شرکتکننده در این نوع انتقال فعال ثانویه دارند، به آنها پروتئینهای «مبادلهکننده» (Exchanger) گفته میشود. برای مثال آنتیپورترهای کلر-بیکربنات در غشای بازولترال سلولهای دیواره معده، یک یون کلر را وارد سیتوپلاسم و یک یون بیکربنات را از سیتوپلاسم خارج میکنند. این پروتئینهای مبادلهکننده در تبادل گاز نقش بسیار مهمی دارند. در غشای گلبولهای قرمز در جریان خون مویرگهای محیطی، با وارد کردن یک را با یک پلاسمای خون مبادله میکند. بی کربنات با واکنش آنزیمی بهوسیله کربونیکانیدراز به گاز تبدیل میشود. اما زمانی که خون در مویرگهای ریه جریان دارد، جهت مبادله این دو یون معکوس میشود. در نتیجه با ورود هر به سیتوپلاسم اریتروسیت، یک با عبور از غشا، از این سلول خارج میشود. گاز دیاکسید کربن حاصل از فعالیت کربونیکانیدراز در نهایت وارد کیسههای هوایی ریه خواهد شد.
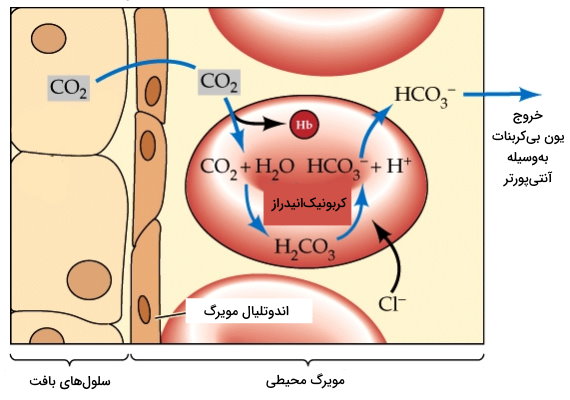
آنتیپورتر یکی دیگر از پروتئینهای عرض غشایی است که را وارد سیتوپلاسم و را از سیتوپلاسم خارج میکند. این ناقلها در بسیاری از بافتها وجود دارند اما تعداد آنها در لوله جمعکننده ادرار و سلولهای اپیتلیال لوله پیچخورده نزدیک بیشتر است. آنژیوتانسیون II یکی از هورمونهایی است که فعالیت آنتیپورتر در نفرونها را تنظیم میکند. اتصال این گیرنده به رسپتورهای سطح غشایی سلولهای اپیتلیال، بازجذب سدیم از نفرون و ترشح یون هیدروژن را افزایش میدهد.
آنتیپورتر (NCX) از اختلاف پتانسیل الکتریکی یون سدیم برای خارج کردن یونهای کلسیم از سیتوپلاسم بهره میبرد. در سلولهای تحریکی (نورون و ماهیچه) این ناقل علاوه بر غشای پلاسمایی در غشای میتوکندری و شبکه اندوپلاسمی صاف، با وارد کردن سه سدیم به سیتوپلاسم، ۱ یون کلسیم را به خارج سلول یا لومن اندامک میفرستد.
سوالات متداول
در این بخش به تعدادی از سوالات متداول پیرامون انتقال فعال سلولی پاسخ میدهیم.
تفاوت انتقال فعال و انتشار تسهیل شده چیست ؟
انتقال فعال و انتشار تسهیل شده دو مکانیسم مختلف برای عبور مولکولها از عرض غشای سلول هستند. در هر دو این فرایندها مولکول پس از برهمکنش با پروتئینهای عرض غشایی وارد سلول یا از آن خارج میشود. تفاوت اصلی این دو در مصرف انرژی و جهت شیب الکتروشیمیایی مواد است. کانالها و ناقلهای پروتئینی انتشار تسهیل شده بدون هیدرولیز ATP و با تغییر کنفورماسیون پروتئین پس از اتصال به لیگاند مولکولهای بزرگ و قطبی را در جهت شیب غلظت شیمیایی و الکتریکی وارد سلول یا از سلول خارج میکنند اما انتقال فعال با استفاده از انرژی شیمیایی ATP، نور خورشید یا پتانسیل الکتریکی یونها، مولکولهای دیگر را از غشا عبور میدهد. به همین دلیل ناقلهای انتشار تسهیل شده تکانتقال یا یونیپورت هستند اما ناقلهای انتقال فعال در هر چرخه عملکردی بیش از یک مولکول را منتقل میکنند.
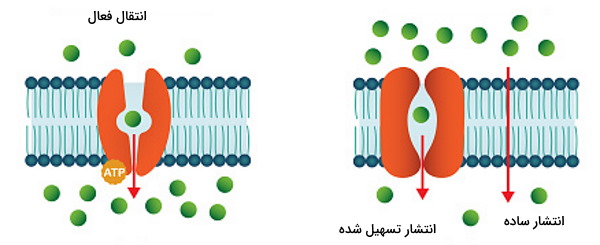
پروتئین های انتقال فعال کدامند ؟
مجموعهای از گلیکوپروتئینهای عرض غشایی در انتقال فعال اولیه و ثانویه شرکت میکنند. پمپها هیدرولازهای ATP و مجموعهای ساختارهای بتاشیت و آلفا هلیکس هیدروفوب و هیدروفیل هستند. تغییر کنفورماسیون این ساختار با هیدرولیز ATP و فسفوریله شدن یک بخش از زنجیره سبب انتقال فعال اولیه از عرض غشا میشوند. همانتقالها (سیمپورتر و آنتیپورتر)، پروتئینهای عرض غشایی هستند که از انرژی الکتروشیمیایی برای تغییر کنفورماسیون ساختار و انتقال فعال ثانویه استفاده میکنند.
تفاوت انتقال فعال و اسمز چیست ؟
اسمز انتقال مولکولهای آب از عرض یک غشای نیمهتروا است که به دلیل اختلاف غلظت ایجاد میشود. در واقع اسمز نوعی انتشار و انتقال غیرفعال است. در این فرایند مولکولهای آب از سمت رقیقتر غشا (مولکول آب بیشتر) به سمت غلیظتر (مولکول آب کمتر) حرکت میکنند و حرکت مولکولها تا به تعادل رسیدن غلظت آب در دو طرف ادامه دارد. اما انتقال فعال نیاز به پروتئینهای غشایی و مصرف انرژی برای انتقال مواد خلاف شیب غلظت دارد.
آیا اندوسیتوز نوعی انتقال فعال است ؟
اندوسیتوز یکی از مسیرهای ورود مواد به سیتوپلاسم سلولهای یوکاریوتی است. در این روش تغییر غشای پلاسمایی سبب ایجاد وزیکولهای غشایی و ورود مواد همراه با بخشی از مایع خارج سلولی میشود. برای تشکیل وزیکولها نیاز به مصرف انرژی است. به همین دلیل از این نظر میتوان اندوسیتوز را نوعی انتقال فعال در نظر گرفت. اما در انتقال فعال اولیه و ثانویه، پروتئینهای عرض غشایی با استفاده از انرژی موجود در ATP یونها و مولکولهایی کوچکتر مثل گلوکز را خلاف جهت شیب غلظت از عرض غشای پلاسمایی عبور میدهند.
فرق انتقال فعال اولیه و ثانویه چیست ؟
انتقال فعال اولیه و ثانویه هر دو از مکانیسمهای تبادل مواد سلولها با محیط هستند که به انرژی نیاز دارند. در انتقال فعال اولیه انرژی لازم برای فرایند از هیدرولیز مولکول ATP تامین میشود. اما انتقال فعال ثانویه اختلاف پتانسیل الکتریکی یک یون بین سیتوپلاسم و ماتریکس خارج سلولی برای انتقال مولکولها (آمینواسید و گلوکز) یا یونهای دیگر استفاده میکند.
جمع بندی
در این مطلب توضیح دادیم که انتقال فعال یکی از مکانیسمهای سلول برای انتقال مواد به و خارج از سیتوپلاسم است. ویژگی که این نوع انتقال مواد را از سایر مکانیسمهای انتقالی متمایز میکند، استفاده از ناقلهای عرض غشایی برای انتقال مواد در جهت خلاف شیب غلظت الکتروشیمیایی است. انرژی لازم برای این انتقال بهوسیله هیدرولیز پیوند فسفودیاستر در ATP (انتقال فعال اولیه) یا اختلاف پتانسیل الکتریکی یک یون (انتقال فعال ثانویه) تامین میشود.










جامع، کامل، مفید، آموزنده و بسیار عالی