



وزیکول چیست؟ – به زبان ساده + انواع، کار و وظیفه
سلولهای یوکاریوتی از اندامکهای مختلفی تشکیل شدهاند که هر کدام وظیفه مشخصی در سلول دارند. برای مثال شبکه اندوپلاسمی زبر، پروتئینهایی را سنتز میکند که برای تغییرات پس از ترجمه به دستگاه گلژی فرستاده میشوند. انتقال مواد بین این اندامکها بهوسیله کیسههای غشایی به نام وزیکول انجام میشود. وزیکولها در پروکاریوتها نیز تشکیل میشوند و علاوه بر انتقال مواد به شناور ماندن موجود روی آب و ذخیره مواد کمک میکنند. در این مطلب توضیح میدهیم وزیکول چیست و انواع مختلف آن چه وظیفهای دارد.
- خواهید آموخت که وزیکول چیست و چگونه در ساختار سلول تشکیل میشود.
- انواع اصلی وزیکولها و تفاوت عملکرد هرکدام را خواهید شناخت.
- نقش وزیکولها در انتقال مواد، ترشح هورمون و ایمنی را یاد میگیرید.
- روشهای جابهجایی وزیکول درون سلول و نقش اسکلت سلولی را یاد خواهید گرفت.
- تفاوت وزیکول با اندامکهای مشابه مانند واکوئل و کاربردهای خاص هر کدام را میآموزید.
- با اهمیت وزیکولها در سلامت سلولی و نقش آنها در بیماریها آشنا میشوید.
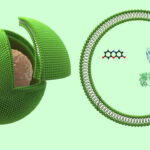



وزیکول چیست ؟
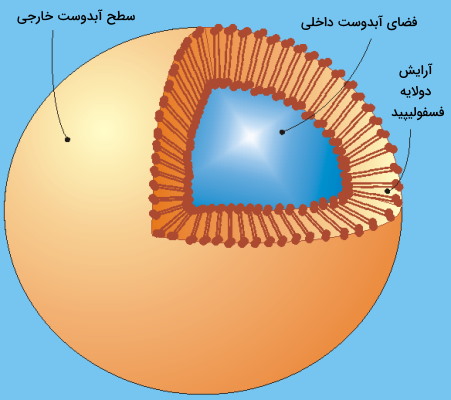
وزیکول ساختارهایی کروی است که در یوکاریوتها مثل غشای پلاسمایی سلول از دو لایه فسفولیپیدی تشکیل میشود. این ساختارها در انواع سلولهای گیاهی، جانوری، باکتریها و آرکیباکتریها تشکیل میشوند و در انتقال بسیاری از مواد بین اندامکهای مختلف یک سلول یا بین سلولهای مختلف شرکت میکنند. از آنجا که وزیکولها معمولا از غشای پلاسمایی سلول یا اندامکهای غشادار داهخل سلول جوانه میزنند، آرایش فسفولیپیدها در آنها مشابه غشای سلولی و بهشکلی است که بخش فسفر قطبی (هیدروفیل) در سطح بیرونی داخلی و لپیدهای هیدروفوب بین آنها قرار میگیرند. لیپوزومها، ساختارهای مشابه وزیکول هستند که در آزمایشگاه سنتز میشوند.
کار وزیکول چیست ؟
وزیکولها ساختارهای ساده درونسلولی هستند که عملکرد بسیار متنوعی دارند. بسیاری از این ساختارها مواد مورد نیاز سلول را ذخیره یا مولکولهای پیامرسان را به خارج سلول منتقل میکنند. این ساختارهای غشایی در فرایندهای مختلف سلولی شرکت میکنند.
- متابولیسم: وزیکولهای خارج سلولی با انتقال مواد از یک سلول به سلول دیگر در متابولیسم نقش دارند.
- نقل و انتقال مواد: وزیکولها مواد را درون سلول، بین سلولهای مختلف و خارج از سلول منتقل میکنند.
- شناور شدن سلول: سیانوباکتریها و هالوباکترهای آبزی به وسیله وزیکولها در آب شناور میمانند.
- ذخیره مواد: آنزیمها، پروتئینها و مولکولهای چربی ازجمله موادی هستند که در وزیکولها ذخیره میشوند.
- ترشح مولکول پیامرسان: ترشح برخی هورمونها از سلولهای اندوکرین و انتقالدهندههای عصبی از نورونها بهوسیله وزیکولها انجام میشود.
- تجزیه مواد زائد: وزیکولها پس از ادغام با لیزوزومها از آنزیمهای این اندامک برای تجزیه مواد مختلف استفاده میکنند.
انواع وزیکول چیست ؟
وزیکولها اجزای مهم سیستمهای انتقالی اندوسیتوز (پینوسیتوز، اندوسیتوز با واسطه گیرنده و فاگوسیتوز)، اگزوسیتوز، انتقال مواد بین اندامکهای یوکاریوت یا سلولهای مختلف هستند. به علاوه انواعی از این ساختارهای کروی در سلولهای مختلف مولکولهای شیمیایی یا گیرندههای مختلف را ذخیره میکنند.
وزیکولها را میتوان بر اساس نوع فعالیتی که انجام میدهند، محل انجام فعالیت و نوع موادی که حمل میکنند به انواع مختلف انتقالی، ترشحی، ذخیرهای و خارج سلولی تقسیمبندی کرد.
وزیکولهای انتقالی
وزیکولهای انتقالی مواد را بین اندامکهای مختلف سیتوپلاسم یا بین دو سلول مختلف منتقل ميکنند. برای مثال پروتئینهای سنتز شده در شبکه اندوپلاسمی زبر بهوسیله این ساختارهای غشایی به دستگاه گلژی و پس از تغییرات به لیزووم، غشای پلاسمایی، هسته یا خود شبکه اندوپلاسمی منتقل میشوند. انتقال این وزیکولها در سلول به مشارکت میکروتوبولها و موتورپروتئینهای اسکلت سلولی نیاز دارد. این وزیکولها به دو دسته پوششدار و بدون پوشش تقسیم میشوند.
وزیکول های پوشش دار
انتقال بسیاری از پروتئینها و لیپیدها بین شبکه اندوپلاسمی، دستگاه گلژی و لیزوزوم به کمک وزیکولهای پوشش دار انجام میشود. این وزیکولها با کمک خودآرایی ( Self-Assembly) بعضی پروتئينها تشکیل میشوند. کلاترین، COPII و COPI سه نوع پروتئينی هستند که در تشکیل وزیکولهای پوششدار شرکت میکنند.
- پوشش کلاترین: وزیکولهای کلاترینی در مسیرهای اندوسیتوز و انتهای مسیرهای ترشحی تشکیل میشوند.
- پوشش COPII: این وزیکولها پروتئین را از شبکه اندوپلاسمی خارج میکنند.
- پوشش COPI: وظیفه این وزیکولها انتقال مواد بین بخشهای مختلف دستگاه گلژی و از دستگاه گلژی به شبکه اندوپلاسمی است.
وزیکول های پوشش دار کلاترین
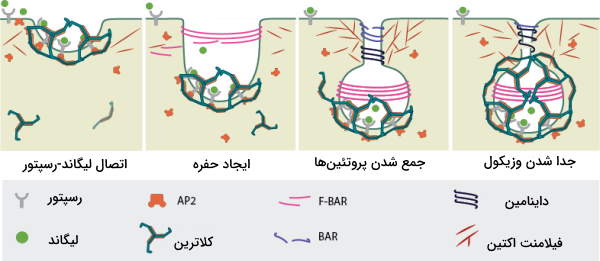
تمام وزیکولهای پوششدار کلاترین با مکانیسم مشابهی تشکیل میشوند. ما برای توضیح این مکانیسم خروج پروتئينها از شبکه اندوپلاسمی زبر (RER) را بررسی میکنیم. پروتئینها پس از تشکیل در شبکه اندوپلاسمی زبر باید برای ایجاد تغییرات پس ترجمه به دستگاه گلژی یا برای ترشح و خارج شدن از سلول به غشای پلاسمایی فرستاده شوند. خروج این پروتئین بهوسیله وزیکولهای پوششدار و در چند مرحله انجام میشود.
- برای شروع، پروتئین یا «بار» (Cargo) به گیرندههای خود (G پروتئین عرض غشایی) در بخش لومنی غشای شبکه اندوپلاسمی متصل میشود.
- پروتئینهای «آداپتین» (Adaptin) به انتهای سیتوپلاسمی رسپتورها متصل میشوند و گیرندهها را کنار هم جمع میکنند.
- اتصال آداپتین به گیرنده سبب تجمع مولکولهای RAB و V-SNARE در سطح سیتوپلاسمی شبکه اندوپلاسمی میشود.
- کلاترین مولکول دیگری است که به این مجموعه اضافه میشود و شبکهای پروتئینی اطراف بخش سیتوپلاسمی شبکه اندوپلاسمی تشکیل میدهد.
- تشکیل شبکه کلاترینی سبب خمیده شدن و شروع جوانه زدن غشا به سمت سیتوپلاسم میشود.
- داینامین (Dynamin) در محل اتصال وزیکول به RER (گردن وزیکول) متصل میشود. داینامین با مصرف GTP دو طرف غشا را به هم متصل و وزیکول را آزاد میکند.
مولکولهای پروتئینی مختلفی در اتصال وزیکول پوششدار به غشای هدف (گلژی و غشای پلاسمایی سلول) شرکت میکنند. T-SNARE و «پروتئینهای متصلکننده» (Tethering Proteins) دو مولکول عرض غشایی در این فرایند هستند.
- پروتئینهای متصلکننده (مولکولهای Sec) با RAB موجود در غشای وزیکول برهمکنش و وزیکول را به غشای میزبان قلاب میکند.
- در مرحله بعد مولکولهای T-SNARE و V-SNARE به هم متصل میشوند و سبب نزدیک شدن وزیکول به غشای هدف و پراکنده شدن کلاترینها از دور وزیکول میشود.
- در نهایت نزدیک شدن این دو غشا به هم منجر به ادغام شدن غشای آنها خواهد شد.
- تفاوت pH اندامک مقصد و مبدا سبب جدا شدن مولکول از گیرنده میشود.
وزیکول های پوشش دار COP
ارتباط بین شبکه اندوپلاسمی و جسم گلژی در سلولهای یوکاریوتی در دو مسیر پیشرونده (شبکه اندوپلاسمی به گلژی) و پسرونده (گلژی به شبکه اندوپلاسمی) انجام میشود. مسیر پیشرونده از وزیکولهای پوششدار COP II و مسیر پسرونده از وزیکولهای پوششدار COP I برای انتقال مواد کمک میگیرند. مکانیسم تشکیل این دو وزیکول با هم متفاوت است.
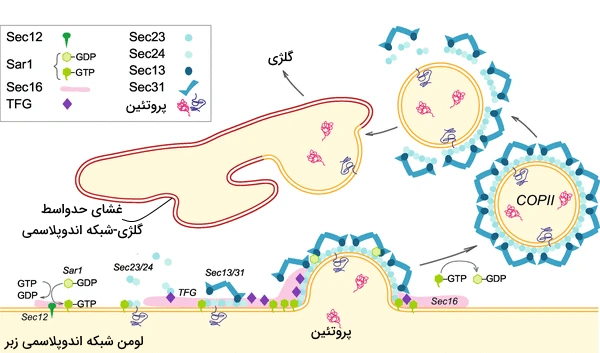
وزیکولهای پوششدار COP II
تشکیل این وزیکولها مثل وزیکولهای کلاترینی وابسته به اتصال مولکول انتقالی با رسپتور داخلی ER است. تشکیل این وزیکولها مثل وزیکولهای کلاترینی و COP II در چند مرحله و به کمک پروتئینهای مختلف انجام میشود.
- GTPase شرکتکننده در تشکیل این وزیکولها Sar1 است. در مرحله اول این پروتئين با غشای ER برهمکنش میکند.
- Sec12 (فاکتور تبادل گوانین) پس از برهمکنش Sar1 با غشای ER مولکول GDP آن را با GTP تعویض و این مولکول را فعال میکند.
- فعال شدن Sar1 سبب ورود بخش آمفیپاتیک این مولکول به غشای ER میشود.
- Sar1 سبب تجمع پروتئين هترودیمر Sec23-Sec24 در اطراف غشای ER خواهد شد. زیرواحد Sec23 به Sar1 و زیرواحد Sec24 به گیرنده مولکول انتقالی متصل میشود.
- تجمع این مجموعه پروتئينی سبب جمع شدن پروتئينهای Sec13/Sec31 در محل تشکیل وزیکول میشود. این پروتئينها پلیمریزه شده و پوشش دوم این وزیکولهای خروجی از ER را تشکیل میدهند.
- هیدرولیز GTP موجود در Sar1 قبل از رسیدن وزیکولها به بخش نزدیک گلژی سبب آزاد شدن پوشش پروتئينی در سیتوپلاسم میشود.
وزیکولهای پوششدار COP I
این وزیکولها مولکولهای مختلف ازجمله لیپیدهای غشای پلاسمایی و V-SNARE را به شبکه اندوپلاسمی برمیگرداند. تشکیل این وزیکولها در چند مرحله انجام میشود.
- (ARF1 (ADP Ribosylation Factor با غشای گلژی برهمکنش میدهد.
- فاکتور انتقال گوانین GDP این مولکول را با GTP تعویض و ARF1 را فعال میکند.
- زنجیره آلفا ARF1 بین لیپیدهای غشای گلژی قرار میگیرد و سبب تجمع مولکولهای COP I اطراف غشای سیتوپلاسمی گلژی میشود.
- COP I شبکهای پروتئينی اطراف غشا تشکیل میدهد که سبب خمیده شدن غشا به فضای سیتوپلاسمی و تشکیل وزیکول خواهد شد.
- قبل از ادغام شدن وزیکول با غشای ER مولکول GTP موجود در ARF1 با GDP مبادله و پوشش COP I در سیتوپلاسم آزاد میشود.
وزیکولهای بدون پوشش
سه دسته وزیکول بدون پوشش در انتقال مواد شرکت میکنند. در بخش قبل توضیح دادیم که وزیکولهای انتقالی کلاترین، COP I و COP II قبل از اتصال به غشای هدف پوشش خود را از دست میدهند. به علاوه فرایند پینوسیتوز یکی از انواع اندوسیتوز است که با استفاده از مولکول ATP بخشی از ماتریکس خارج سلولی و مولکولهای محلول در آن را بهوسیله وزیکولهای بدون پوشش وارد سلول میکند. برای تشکیل این وزیکولها نیاز نیست مولکول به گیرنده سطح غشایی متصل شود. این وزیکولها بسیار کوچکتر از وزیکولهای پوششدار اندوسیتوزی، فاگوزومها و اتوفاژها هستند. مولکولهای چربی و ویتامینها به کمک این وزیکولها وارد سلولهای دیواره روده باریک و پس از ورود به لیزوزوم تجزیه میشوند. وزیکولهای فرایند اگزوسیتوز (وزیکولهای ترشحی) نوع دیگری از وزیکولهای بدون پوشش هستند.
فاگوزومها
این وزیکولها ازجمله وزیکولهای بدون پوششی هستند که در فرایند فاگوسیتوز مواد زائدی که باید در سلول جانوری تجزیه شود را به لیزوزوم منتقل میکنند. این وزیکولها در ماکروفاژها، نوتروفیلها و سلولهای دندریتی (سلولهای بیگانه خوار سیستم ایمنی) تشکیل میشوند. بعضی از باکتریها از فاگوزومها برای ورود به سلول میزبان استفاده میکنند. تقسیم دوتایی و رشد این باکتریها (کوکسیلا) در لیزوزوم سلول جانوری انجام میشود یا قبل از ادغام فاگوزوم با لیزوزوم باکتری وارد سیتوپلاسم خواهند شد (ریکتسیا). در جانوران تکسلولی ازجمله آمیب مواد غذایی مورد نیاز جاندار در این وزیکولها قرار دارد.
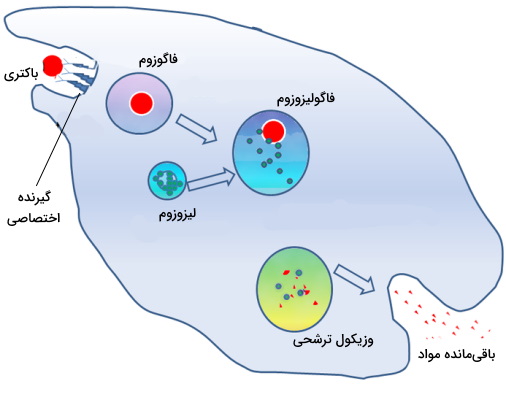
اتوفاگوزوم ها
اتوفاگوزومها گروهی از وزیکولهای درونسلولی هستند که پروتئینهای غیرطبیعی سیتوپلاسم و اندامکهای اضافه یا تجزیه شده را برای تجزیه در فرایند اتوفاژی به لیزوزوم منتقل میکنند. تشکیل این وزیکولها بهوسیله ژنهای محافظت شده از مخمرها تا یوکاریوتهای پیشرفته تنظیم میشود. اتوفاگوزومها نزدیک بخشی از شبکه اندوپلاسمی تشکیل میشود که آنزیمهای فسفاتیدیل سنتتاز و ترانسفراز سنتز میشوند. این وزیکولها پس از ادغام با لیزوزوم از آنزیمهای این اندامک برای تجزیه باقیماندههای سلولی و مواد زائد استفاده میکنند.
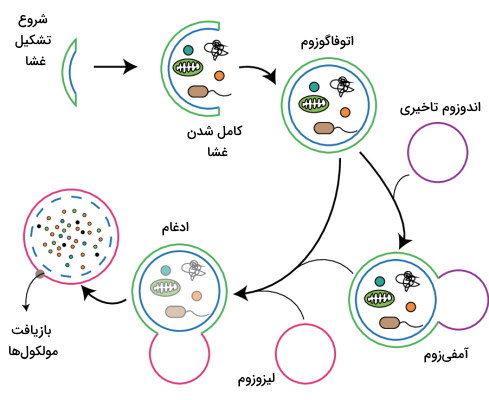
اندازه این وزیکولها بین گونههای مختلف متفاوت و از ۵۰۰ تا ۱۵۰۰ نانومتر تغییر میکند. در سلولهای بنیادی جنینی، فیبروبلاستهای جنینی و سلولهای کبدی فاگوزومها را میتوان بهوسیله میکروسکوپ نوری مشاهده کرد. اختلال در فرایند اتوفاژی و عملکرد اتوفاگوزومها منجر به ایجاد بیماریهای مختلف ازجمله انواع سرطان، بیماریهای تحلیل نورونها و بیماریهای سیستم ایمنی میشود.
نقش اتوفاگوزوم در نورون ها
اتوفاژی یکی از فرایندهای مهم در سلول برای تجزیه پروتئینهایی است که ساختار عملکردی آنها درست شکل نگرفته است. تجمع این پروتئینها در نورون منجر به بسیاری از بیماریهای تحلیل اعصاب میشود.به همین دلیل سرعت اتوفاژی در نورون و تعداد اتوفاگوزومهایی که در این سلول تشکیل میشود نسبت به سایر سلولها بسیار بیشتر است. اختلال در فرایند اتوفاژی نورون منجر به تجمع بقایای اندامکها و پروتئینهای غیرعملکردی در آکسون و دندریت نورون و تورم و تغییر شکل این زوائد سیتوپلاسمی خواهد شد. به علاوه تشکیل این وزیکولها با کنترل رهایش کلسیم از شبکه اندوپلاسمی آکسون در تنظیم سیناپس نقش دارند. اتوفاگوزومها در پایانه آکسون نورونها تشکیل میشوند و برای ادغام با لیزوزوم بهوسیله میکروتوبولها و کاینزین مسیر پسرونده به جسم یاختهای نورون منتقل میشوند.
اندوزوم
اندوزومها وزیکولهای درونسلولی و اولین مقصد وزیکولهایی هستند که بهوسیله اندوسیتوز وارد سلول میشوند. در اندوسیتوز غشای پلاسمایی به داخل سیتوپلاسم تا میخورد، اطراف مولکولی که باید وارد سلول شود را میگیرد و مولکول به همراه بخشی از ماتریکس خارج سلولی وارد سیتوپلاسم میشود. سه نوع اندوزوم در سلولهای یوکاریوتی تشکیل میشود.
اندوزوم اولیه
«اندوزوم اولیه» (Early endosomes) اولین ساختار غشایی است که وزیکولهای اندوسیتوزی ادغام میشود و نزدیک غشای پلاسمایی قرار دارند. مولکول شاخص غشای این اندوزوم فسفااینوزیتید فسفاتیدیل اینوزیتول ۳-فسفات است. به علاوه پمپ پروتونی (V-ATPase) غشای این وزیکول منجر به کاهش pH (۶٫۲) داخلی آن میشود. در بسیاری از سلولها اندوزوم اولیه از یک بخش لولهای (۶۰ nm) و یک حفره (۴۰۰ nm) تشکیل شده است. مهمترین وظیفه این وزیکولها طبقهبندی مواد ورودی به مواد بازیافتی و تجزیهای است. رسپتور همراه مولکولها در بخش لولهای جمع میشوند و پس از طبقهبندی برای شرکت در اندوسیتوز جدید به غشای سلولی برمیگردند. پروتئینها و لیپیدهای غشای پلاسمایی، ازجمله ترانسفرین مولکولهایی هستند که در این بخش جمع میشوند. مولکول ورودی در حفره جمع میشود و پس از طبقهبندی به اندوزوم تاخیری منتقل میشود.
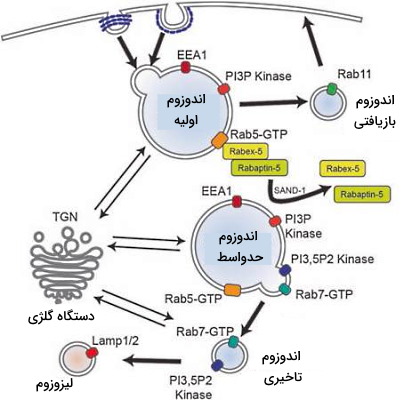
یکی پروتئینهای نشانه اندوزوم اولیه که بهوسیله آن میتوان این وزیکول را از سایر وزیکولها تشخیص داد، پروتئین RAB5 در غشای این وزیکول است که موفولوژی و عملکرد این ساختار غشایی را تنظیم میکند. RAB5 یک G پروتئین کوچک است و با همکاری مولکولهای تبادل GTP اتصال وزیکول اندوسیتوزی به اندوزوم اولیه و فعال شدن مسیر پیامرسانی وابسته به فسفاتیدیل اینوزیتیل ۳-فسفات (IP3) را تنظیم میکند.
اندوزومهای تاخیری یا اجسام چندحفرهای
اندوزومهای تاخیری یا اجسام چند حفرهای (Multivesicular bodies or late endosomes) وزیکولهای نزدیک دستگاه گلژی هستند. این اندوزومها از بالغ شدن یا تغییر اندزومهای اولیه به وجود میآیند. وزیکولهای تاخیری یا با لیزوزومهای قبلی سلول ادغام میشوند یا با دریافت آنزیمهای هیدرولازی تبدیل به لیزوزوم جدید خواهند شد. یکی از نشانههای تبدیل شدن اندوزوم اولیه به اندوزوم تاخیری جایزینی G- پروتئين Rab5 به Rab7 است. غشای این وزیکولها از یک لیپید اختصاصی به نام BMP یا LBPA تشکیل شده است که در سایر غشاهای یوکاریوتی وجود ندارد.
اندوزومهای بازیافتی
«اندوزومهای بازیافتی» (Recycling endosomes) این اندوزومها فاصله کمی از غشای پلاسمایی قرار دارند و ترکیبات را به بخش ترانس دستگاه گلژی یا غشای پلاسمایی انتقال میدهند. تعداد این اندوزومها در مرکز سازماندهی توبولها بسیار زیاد است. این وزیکولها در دو مسیر بازیافتی سریع و آهسته شرکت میکنند.
- مسیر بازیافتی سریع: در این مسیر موادی بازیافتی ازجمله رسپتورهای اندوسیتوز مستقیم از اندوزوم اولیه به غشای پلاسمایی برمیگردند. برای تشکیل وزیکولهای این مسیر به Rab4 و Rab35 نیاز است.
- مسیر بازیافتی آهسته: وزیکولهای این مسیر بهوسیله Rab11a تنظیم میشوند. در این مسیر وزیکولها قبل از غشای پلاسمایی وارد دستگاه گلژی میشوند.
وزیکول های ذخیره ای
وزیکولهای ذخیرهای ساختارهای ویژه سیتوپلاسم سلولهای ماهیچهای و چربی هستند که GLUT4 (Glucose Transporter Type 4) ذخیره میکنند. اتصال انسولین به گیرنده خود در سطح این سلولها با ادغام این وزیکولها با غشای سلول و اگزوسیتوز ناقل GLUT4 همراه است.
گلیزومها
گلیزومها وزیکولهای ذخیرهای در یاخته های پشتیبان (سلولهای گلیا) سیستم عصبی مرکزی هستند که آنزیمهای لازم برای تقسیم سلولی و مولکولهای تنظیمکننده سیناپس را ذخیره میکنند. گلیزومها در یاختههای پشتیبان آستروسیت، حاوی پروتئينهای شرکتکننده در فرایند اگزوسیتوز آستروسیتها (VAMP3)، پروتئينهای اسکلت سلولی آستروسیت پیشسیناپسی (ازرین | Ezrin) و گلیکوپروتئینهای غشای آستروسیت (باسیگین | Basigin) و G-پروتئینهای مسیر انتقال پیام است.
وزیکولهای ترشحی
وزیکولهای ترشحی در یوکاریوتها از دستگاه گلژی، شبکه اندوپلاسمی زبر و شبکه اندوپلاسمی صاف جوانه میزنند و پروتئینها یا لیپیدهای سنتز شده در این اندامکها را از سلول خارج میکنند. بعضی وزیکولهای ترشحی ازجمله وزیکولهای غشایی دیواره میزنای، وزیکولهای اندوسیتوزی هستند که مستقیم از سلول خارج میشوند. دو مسیر خروجی برای وزیکولهای ترشحی وجود دارد.
- ترشح یا اگزوسیتوز ثابت: این نوع اگزوسیتوز در تمام سلولهای یوکاریوتی وجود دارد که پروتئینها و لیپیدهای غشایی سنتز شده در شبکه اندوپلاسمی زبر، دستگاه گلژی و شبکه اندوپلاسمی صاف را به بخش خارجی غشا منتقل میکند.
- ترشح یا اگزوسیتوز تنظیم شده: این مسیر در سلولهای ترشحکننده هورمون (اندوکرین اگزوکرین)، انتقالدهندههای عصبی و آنزیمهای گوارشی وجود دارد و بهوسیله مولکولهای پیامرسان تنظیم میشود. این وزیکولها برخلاف وزیکولهای اندوسیتوزی نیازی به پوشش پروتئینی برای انتقال ندارند. غشای این وزیکولها به شکل دائمی با غشای پلاسمایی ادغام نمیشود و پس از خارج شدن مواد، اندوسیتوز آنها را به سیتوپلاسم برمیگرداند.
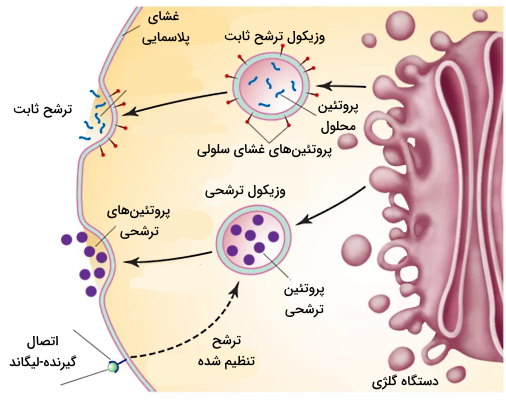
اندازه این وزیکولهای ترشحی سلولهای اگزوکرین که هورمونهای خود را به مجرای غدد میریزند، از همه بیشتر (۵۰ نانومتر) و اندازه وزیکولهای سیناپسی نورون از همه کمتر (1 میکرومتر) است. این وزیکولها بر اساس ویسکوزیتی مایع درون غشا به دو دسته تقسیم میشوند.
- وزیکولهای سروزی: مایع درون این وزیکولها ویسکوزیتی کمی دارد. بیشتر وزیکولهای سروزی آنزیمهای غیرفعال را از سلول خارج میکنند. به دلیل غلظت بالای پروتئین در این وزیکولها که سبب افزایش چگالی الکترون میشود، به راحتی میتوان این وزیکولها را با میکروسکوپ الکترونی مشاهده کرد. این وزیکولها ترشحات غدد پاراتیروئید، غدد اشکی، برخی آنزیمهای پانکراس و غدد عرق را از سلول خارج میکنند.
- وزیکولهای موکوزی: مابع درون این وزیکولها به دلیل غلظت زیاد موسین اسیدی (ترکیب گلیکوپروتئین و گلیکوساکارید)، ویسکوزیته زیادی دارد. این وزیکولها را میتوان بهوسیله رنگآمیزی پرییدیکاسید شفت (PAS) و با میکروسکوپ نوری مشاهده کرد. این وزیکولها ترشحات سلولهای جامیشکل در اپتلیال معده را از سلول خارج میکنند.
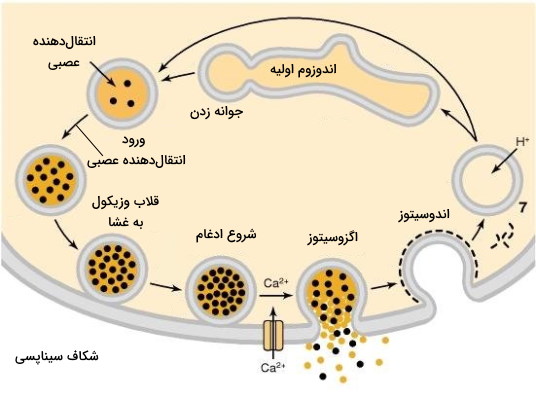
وزیکول های سیناپسی
وزیکولهای سیناپسی محل ذخیره انتقالدهندههای عصبی غیرپروتئینی هستند که پس از رسیدن پیام عصبی به پایانه آکسون با غشای پایانه ادغام میشوند و نروترسمیترها را به فضای سیناپسی ترشح میکنند. به همین دلیل میتوان این وزیکولها را در دو دسته ذخیرهای و ترشحی قرار داد. وزیکولهای سیناپسی مانند سایر وزیکولها از ترکیب لیپید و پروتئین تشکیل میشوند. فسفاتیدیل کولین، فسفاتیدیل سرین، فسفاتیدیل اینوزیتول و کلسترول لیپیدهای تشکیل دهنده این وزیکولها هستند. پروتئینها سهم کمی در ساختار وزیکولها دارند.
V-ATPase ناقل پروتون موجود در این وزیکولها است که با ایجاد اختلاف غلظت هیدروژن، ورود انتقالدهنده عصبی به وزیکول را تسهیل میکند. به علاوه هر انتقالدهنده عصبی ناقل اختصاصی خود را دارد. برای مثال پروتئین غشایی VAChT (Vesicular Acetylcholine Transporter) در غشای وزیکولهایی وجود دارد که انتقالدهنده عصبی استیل کولین را ذخیره و به شکاف سیناپسی ترشح میکنند. همچنین پروتئينهای SNARE در غشای این وزیکولها به انتقال و ادغام شدن آنها با غشای پایانه آکسون کمک میکنند. اگزوسیتوز وزیکولهای سیناپسی در چند مرحله انجام یشود.
- انتقالدهنده عصبی بهوسیله انتقال فعال و مصرف ATP وارد وزیکول میشود.
- وزیکول بهوسیله موتورپروتئینها و میکروتوبولهای موجود در پایانه به غشای آکسون منتقل میشود.
- وزیکول به غشای پایانه آکسون نورون پیشسیناپسی قلاب و ادغام دو غشا (با مصرف ATP) شروع میشود.
- باز شدن کانالهای ولتاژی کلسیم سبب ورود یون کلسیم به پایانه آکسون و ادغام وزیکول با غشا و انتقالدهنده عصبی وارد شکاف سیناپسی خواهد شد.
- وزیکول ادغام شده با غشا بهوسیله پروتئینهای کلاترین پوششدار و با ورود یون هیدروژن اسیدی میشود.
- در مرحله آخر این وزیکول با اندوزوم اولیه ادغام شده و پس از آماده شدن جوانه میزند و چرخه را ادامه میدهد. بعضی وزریکولها وارد این مرحله نمیشوند.
وزیکول های ترشحی هورمون
هورمون انسولین و گلوکاگون، دو هورمونی هستند که بهوسیله سلولهای بتای پانکراس برای تنظیم قند خون تولید و ترشح میشوند. با افزایش غلظت گلوکز در خون، وریکولهایی که انسولین ذخیره کردهاند با غشای سلول ادغام و انسولین وارد خون میشود. اثر گلوکاگون عکس انسولین است و در موارد کاهش گلوکز خون، ترشح آن منجر به تجزیه گلیکوژن به گلوکز میشود و قند خون را افزایش میدهد. به علاوه وزیکولهای ترشحی پانکراس وظیفه انتقال آنزیمهای گوارشی (پروتئازها، لیپازها و آمیلازها) به مجرای مشترک را بر عهده دارند. هورمون رشد، هورمون تحریککننده فولیکول (FSH)، اکسیتوسین، پرولاکتین و هورمون ضدادراری (ACTH) سایر هورمونهایی هستند که بهوسیله این مکانسیم ترشح میشوند.
وزیکولهای خارج سلولی
وزیکولهای خارج سلولی یا اگزوزومها از اندوزومها جوانه میزنند و وظیفه انتقال مولکولهای زیستی مختلف ازجمله پروتئین، لیپید و اسیدنوکلئیک بین سلولها را بر عهده دارند. اگزوزومها در بعضی انواع سرطان، بیماریهای تحلیل سیستم عصبی و بیماریهای التهابی، عوامل بیماریزا را منتقل میکنند. برای مثال در بعضی متاستازهای سرطان پستان، اگزوزومهای حاوری mRNA ژن سرطان در مایعات بدن افزایش مییابد.
نقش اسکلت سلولی در انتقال وزیکولها
میکروفیلامنتهای اکتین و میکروتوبولها دو عضو اصلی اسکلت سلولی هستند که علاوه بر تعیین و حفظ پایداری ساختار سلول در انتقال وزیکولهای درونسلولی نقش دارند. بررسی نقش این رشتههای پروتئین در انتقال وزیکولها برای اولین بار در سلولهای کرومافین (سلولهای ترشحکننده اپینفرین و نوراپینفرین در فوق کلیه) و بهوسیله مهارکنندههای رشتههای پروتئينی و میکروسکوب کانفوکال انجام شد. نتایج حاصل از این این مطالعات دو حقیقت در مورد نقش اکتین و میکروتوبول در انتقال وزیکولهای درونسلولی را آشکار کرد.
- مهارکنندههای میکروتوبول حرکت وزیکولهای اطراف هسته و مهارکنندههای اکتین حرکت وزیکولها نزدیک هسته و نزدیک غشای سلولی را مختل میکند. در نتیجه میتوان گفت میکروتوبولها بیشتر در مراحل ابتدایی انتقال وزیکولهای ترشحی و میکروفیلامنتها در مراحل پایانی ترشح سلولهای اندوکرین نقش دارند.
- مهارکنندههای اکتین مسیر انتقال سریع و آهسته وزیکولها را مختل میکنند اما مهارکنندههای میکروتوبولها تنها مسیر آهسته ترشح را مختل میکنند.
میوزین، موتورپروتئین همراه میکروفیلامنتهای اکتین است که با هیدرولیز ATP انرژی لازم برای انتقال وزیکولها را فراهم میکند. این موتورپروتئینها از یک بخش اتصالی به وزیکول، یک بخش اتصالی به اکتین و رابط این دو بخش تشکیل میشوند. ۳ نوع میوزین نیروی لازم برا انتقال وزیکولها در مسیرهای مختلف را تامین میکنند.
- میوزین نوع I و VI: در انتقال وزیکولهای اندوسیتوزی به بخشهای درونی سیتوپلاسم نقش دارند.
- میوزین نوع V: در انتقال وزیکول بین اندامکها نقش دارد.
میکروتوبولها رشتههای پروتئينی پویا در اسکلت سلولی هستند که یک انتهای مثبت (تشکیل) و یک انتهای منفی (تخریب) دارند. این پروتئینها به کمک موتورپروتئینهای کاینزین (حرکت به سمت انتهای مثبت) و داینئین و با مصرف ATP وزیکولها را بین اندامکهای مختلف حرکت میدهند. در انتقال وزیکولهای از جسم سلولی نورون به پایانه آکسون و مخالف آن، میکروتوبولها نقش اصلی را دارند. برای مثال در حرکت وزیکولها و میتوکندری از جسم سلولی نورون به پایانه آکسون وزیکولها به کمک کاینزین و در انتقال وزیکولها از پایانه آکسون به جسم سلولی وزیکولها به کمک داینئین منتقل میشوند.
تفاوت واکوئل و وزیکول چیست ؟
واکوئلها و وزیکولها هر دو ساختارهای غشایی درونسلولی هستند. در بعضی تقسیمبندیها واکوئل را نوعی وزیکول در نظر میگیرند. واکوئلها در سلولهای جانوری تشکیل نمیشوند و اندامکهای مخصوص سلول گیاهی، باکتری و قارچها است. واکوئلها بزرگتر از وزیکولها هستند. تفاوت دیگر این دو ساختار در آنزیمهای هیدرولازی است که واکوئل برای تجزیه مواد گیاهی استفاده میکند. وزیکولها با لیزوزوم سلول جانوری ادغام میشوند و بهوسیله آنزیمهای آن مولکولهای ورودی را تجزیه میکنند.
وزیکول های گازی
در بخشهای قبل اشاره کردیم که در بعضی پروکاریوتهای آبزی وزیکولها به شناور ماندن موجود در آب کمک میکنند. وزیکولهای گازی ساختارهای دوکیشکلی هستند که پلانکتونها و سیانوباکتریها را در سطح آب نگه میدارند. این وزیکولها بر خلاف وزیکولهای یوکاریوتی از پروتئینهای آبگریزی تشکیل شدهاند و مولکولهای کربوهیدرات یا لیپید در ساختار آنها وجود ندارد. پروتئین تشکیلدهنده وزیکولهای گازی نسبت به آب نفوذ ناپذیر است اما مولکولهای کوچک گاز از آن عبور میکنند. فشار محیط عامل مهمی در شکل و قطر وزیکولهای گازی است. قطر وزیکولها در گونههای مختلف متفاوت است و هرچه قطر وزیکول بیشتر باشد گاز بیشتری ذخیره میکند. در نتیجه وزیکولها در گونههایی که رشد سریعتری دارند بزرگتر است و این اورگانیسمها را نزدیک سطح آب شناور میکند.
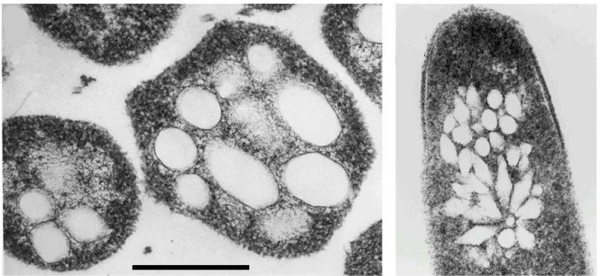
GvpA و GvpC دو پروتئين اصلی وزیکولهای گازی سیانوباکتریها هستند. پروتئين A بسیار آبگریز و تعداد آن در ساختار وزیکول بیشتر است و چارچوب کلی شکل وزیکول را تعیین میکند. پروتئین آبدوست C حفظ تعادل و پایداری وزیکول را بر عهده دارد. تنظیم بیان ژن و عوامل محیطی تشکیل وزیکولهای گازی در سیانوباکتریها را کنترل میکنند. شدت نور، غلظت کربوهیدراتها، غلظت اکسیژن، pH و امواج فراصوت، عوامل محیطی هستند که با تغییر بیان پروتئینها یا به هم ریختن ساختار وزیکول، تشکیل وزیکول گازی را تنظیم میکنند.
سوالات متداول
در این بخش به تعدادی از سوالات متداول پیرامون وزیکول پاسخ میدهیم.
وزیکول سمینال چیست ؟
وزیکول سمینال یکی از غدد دستگاه تولید مثلی مردان است که پشت مثانه و جلوی رکتوم قرار دارد. این غده از از بافت ماهیچهای و سلولهای اندوکرین تشکیل میشود. سلولهای اندوکرینی این غده بخشی از مایع سِمِن را میسازنند که به حرکت اسپرم در مجاری خروجی بیضه کمک میکند. این مایع از فروکتوز، پروستوگلاندینها، ترکیبات بازی و فاکتورهای انعقادی تشکیل شده است. انقباض بافت ماهیچه سبب ورود سمن به میزنای میشود.
وزیکول پوستی چیست ؟
وزیکولهای پوستی یا تاول کیسه کوچک پر از مایعی در سطح پوست هستند که به دلیل آسیب بافت پوست به وجود میآیند. این وزیکولها علاوه بر آسیب مستقیم به لایه خارجی پوست مثل مواردی سوختگی، خراش عفونت باکتریایی یا ویروسی، بر اثر بعضی بیماریهای خودایمنی ایجاد میشوند. درمان این وزیکولها بر اساس عاملی که باعث آن شده، متفاوت است.
وزیکول ژرمینال چیست ؟
وزیکول ژرمینال، هسته سلول تخمکی است که در پروفاز I میوز قرار دارد. پوشش این هسته در پاسخ به LH (هورمون مرحله لوتئال) و قبل از تخمکگذاری از بین میرود. در میوز تخمک ادامه پیدا میکند.
جمعبندی
در این مطلب توضیح دادیم وزیکولها ساختارهای کوچکی هستند که از غشای دولایه لیپیدی یا پروتئینهای آبگریز در یوکاریوت و پروکاریوتها تشکیل میشوند و وظیفه اصلی آنها انتقال مواد بین اندامکهای درونسلولی یا بین سلولهای مختلف است. انواع مختلف این ساختارها در ورود مواد شیمیایی و پاتوژنها به سلول یوکاریوتی نقش دارند.









