


تقسیم سلولی چیست؟ – شرح مراحل به زبان ساده
تقسیم سلولی فرایندی است که در طی آن از یک سلول، دو سلول دختری ایجاد میشود. تقسیم سلولی به دلایل زیادی در موجودات مختلف صورت میگیرد. به عنوان مثال، هنگامی که پوست زانوی شما کنده میشود، سلولهای پوستی برای جایگزینی سلولهای قدیمی، مرده یا آسیبدیده تقسیم میشوند. در این مطلب از مجله فرادرس به بررسی انواع تقسیم سلولی، مراحل آن و چرخه سلولی میپردازیم. در کتابهای علوم پایههای چهارم، ششم، هفتم و هشتم تقسیمات سلولی و انواع آنها توضیح داده شده است که این مطلب میتواند به فهم هرچه بهتر دانشآموزان کمک کرده و اطلاعات مفید دیگری را به دانش آنها اضافه کند.
- یاد میگیرید تقسیم سلولی چیست و چرا رخ میدهد.
- متوجه اهمیت تقسیم سلولی میشوید.
- انواع تقسیم سلولی یعنی تقسیم دوتایی باکتریها و تقسیمهای میوز و میتوز را میشناسید.
- مراحل چرخه سلولی و ورود سلولها به فاز تقسیم هسته را یاد میگیرید.
- سیتوکینز یا روند تقسیم سیتوپلاسم را بررسی میکنید.
- دلایل نوترکیبی در میوز که باعث تولید گامتهای متفاوت میشوند را یاد میگیرید.


تقسیم سلولی چیست؟
تقسیم سلولی فرایندی است که در آن سلول والد تقسیم میشود و در نهایت سلولهای دختر جدیدی را به وجود میآورد. در موجودات تکسلولی، تقسیم سلولی وسیله تولیدمثل است. در موجودات چندسلولی وسیله رشد و نگهداری بافت است در طول زندگی یک فرد، تقسیم سلولی میتوز ادامه مییابد.
در طی تقسیم سلول اطلاعات ژنتیکی آن باید به سلولهای دختری منتقل شوند، زیرا سلولهای دختری بدون محتویات ژنتیکی نمیتوانند چرخه سلولی خود را کامل کنند. مطالعه ساختار کروموزومی، محل و عملکرد آن در سلولها یکی از زیرشاخههای ژنتیک به نام «سیتوژنتیک» را ساخته است که برای یادگیری کامل آن پیشنهاد میدهیم از فیلم آموزش سیتوژنتیک فرادرس استفاده کنید که لینک دسترسی به آن در کادر زیر درج شده است.
در برخی از بافتها مانند اپیتلیوم (پوست، غشاهای مخاطی)، میتوز به طور فعال برای جایگزینی سلولها و ترمیم آسیب رخ میدهد. تقسیم سلولی معمولاً به عنوان بخشی از یک چرخه سلولی بزرگتر رخ میدهد. در یوکاریوتها، دو نوع متمایز از تقسیم سلولی وجود دارد. یک تقسیم میتوز، که به موجب آن هر سلول دختر از نظر ژنتیکی با سلول مادر یکسان است و یک تقسیم سلولی میوز، که به موجب آن تعداد کروموزومها در سلولهای دختر به نصف کاهش مییابد تا گامتهای هاپلوئید تولید شود.

چرا تقسیم سلولی رخ می دهد ؟
آیا تا به حال به نحوه رشد و چرایی رشد گیاهان، حیوانات و حتی انسانها فکر کردهاید؟ پاسخ به دلیل تقسیم سلولی است. اما این فرایند به دلایل دیگری نیز برای موجودات زنده مهم است. در حال حاضر در بدن شما سلولها هر ثانیه در حال تقسیم شدن هستند. تریلیونها سلولی که بدن شما را میسازند، همه از یک سلول به وجود آمدهاند و دلیل اینکه شما اکنون یک ارگانیسم چندسلولی هستید، همه به لطف تقسیم سلولی است. همه موجودات چندسلولی و برخی از موجودات تکسلولی از سلولهای یوکاریوتی تشکیل شده اند.

سلولهای یوکاریوتی به هر نوع سلولی گفته میشود که در آن DNA داخل یک هسته قرار دارد. فرایند تقسیم سلولهای یوکاریوتی برای ایجاد کپیهای یکسان از خود، میتوز نامیده میشود. پروکاریوتها موجوداتی هستند که از یک سلول پروکاریوتی تشکیل شدهاند. سلولهای پروکاریوتی همچنین اطلاعات ژنتیکی مانند DNA دارند، اما این اطلاعات در هسته وجود ندارد. در ادامه دلایل تقسیم سلولی در موجودات را بررسی کردهایم.
رشد
بقای یوکاریوتها به تعامل بین بسیاری از انواع سلول بستگی دارد و ضروری است که توزیع متعادل انواع سلولها حفظ شود. این امر با فرایند بسیار تنظیمشده تکثیر سلولی به دست میآید. رشد و تقسیم جمعیتهای سلولی مختلف به روشهای مختلفی تنظیم میشود، اما مکانیسمهای اساسی در سراسر موجودات چندسلولی مشابه است. بیشتر بافتهای بدن با افزایش تعداد سلولهای خود رشد میکنند، اما این رشد برای حفظ تعادل بین بافتهای مختلف، تنظیم میشود.

در بزرگسالان، بیشتر تقسیم سلولی به جای رشد، در بازسازی بافت نقش دارد، بسیاری از انواع سلولها تحت جایگزینی مداوم هستند. به عنوان مثال، سلولهای پوست به طور مداوم در حال جدا شدن و جایگزینی هستند. در این حالت، سلولهای تمایزیافته بالغ تقسیم نمیشوند، اما جمعیت آنها با تقسیم سلولهای بنیادی نابالغ تجدید میشود. در برخی دیگر از سلولها، مانند سلولهای کبد، سلولهای بالغ قادر به تقسیم باقی میمانند تا اجازه رشد یا بازسازی پس از آسیب را بدهند.
در مقابل این الگوها، انواع دیگر سلولها یا نمیتوانند تقسیم شوند یا توسط مولکولهای زیستی خاصی که توسط سلولهای مجاور تولید میشوند، از تقسیم آنها جلوگیری میشود. در نتیجه، در ارگانیسم بالغ، برخی از بافتها ظرفیت بسیار کاهش یافتهای برای تجدید سلولهای آسیبدیده یا بیمار دارند. نگهداری و تعمیر این سلولها به جای جایگزینی کل سلولها به جایگزینی اجزای داخل سلولی محدود میشود.
تولید مثل
سلولها نیز به عنوان نوعی از تولیدمثل تقسیم میشوند. موجودات تکسلولی مانند باکتریها و آمیبها از تقسیم سلولی برای ساختن کپی از خود استفاده میکنند. در این موارد، فرزندان با والدین یکسان هستند. یک سلول یوکاریوتی برای تولیدمثل از طریق میتوز وارد عمل میشود، در حالی که یک سلول پروکاریوتی فرایندی به نام شکافت دوتایی را طی میکند که در ادامه مطلب مورد بحث قرار خواهد گرفت.
تقسیم سلولی در ارگانیسمهای دارای تولیدمثل جنسی نیز راهی است که این ارگانیسمها گامتها را ایجاد میکنند که سلولهای جنسی (تخمک و اسپرم) محسوب میشوند. این کار از طریق شکل منحصربهفردی از تقسیم سلولی به نام تقسیم میوز انجام میشود که سلولهای دختری منحصربهفرد از لحاظ ژنتیکی را ایجاد میکند.

ترمیم
به آخرین باری که زانوی خود را خراش دادید یا کاغذ بریدید فکر کنید. چرا آن زخم در نهایت خوب شد؟ پاسخ، تقسیم سلولی است. راهی که موجودات زنده پس از آسیبدیدگی بهبود مییابند از طریق تقسیم سلولی است. سلولهای ناحیه آسیبدیده برای جایگزینی سلولهای مرده یا آسیبدیده از طریق میتوز عمل میکنند که به ارگانیسم اجازه میدهد تا بهبود یابد.
سلولها فقط برای مدتی زندگی میکنند. در نهایت سلولهای قدیمی ممکن است به درستی کار نکنند و نیاز به تعویض داشته باشند. هنگامی که یک سلول میمیرد یا آسیب میبیند، سایر سلولهای سالم تقسیم میشوند تا مطمئن شوند که اختلالی در هومئوستاز وجود ندارد. این همیشه اتفاق میافتد تا مطمئن شود که شما و هر موجود زنده دیگری در این سیاره میتوانید به انجام عملکردهای زندگی خود ادامه دهید. تقسیم سلولی در هر نقطه از چرخه زندگی یک موجود زنده تا روز مرگ آنها انجام میگیرد.
اهمیت تقسیم سلول ها چیست؟
تقسیم سلولی یکی از مهمترین فرایندهایی است که در هر موجود زندهای اتفاق میافتد. هنگامی که رشد یک موجود زنده به پایان میرسد، تقسیم سلولی کند میشود اما هرگز متوقف نمیشود. سلولهای قدیمی باید دائما با سلولهای سالم جدید جایگزین شوند. اگر این اتفاق نیفتد میتواند منجر به عوارضی در یک ارگانیسم شود. به عنوان مثال، اگر سلولهای ریه فردی در اثر تنفس آلودگی آسیب ببینند، بدن آن سلولهای آسیبدیده را با سلولهای سالم جایگزین میکند. به همین دلیل است که افرادی که در اثر سیگار کشیدن به ریههای خود آسیب رساندهاند، اگر به سلولهای ریه خود فرصت دهند تا خود را از طریق تقسیم سلولی ترمیم کنند، باز هم میتوانند بهبود یابند.

اما بدون تقسیم سلولی، مشکلات بزرگتری به وجود میآید. کل سیستمهای اندامها به میتوز وابسته هستند تا مطمئن شوند که هر عضوی سالم است و به درستی کار میکند. برای مثال، اگر تمام ماهیچهها و رگهای خونی شما در حالت کار هستند، اما سلولهای سازنده قلب شما به درستی تقسیم نمیشوند تا مطمئن شوید که اندام میتواند به پمپاژ خون در بدن ادامه دهد، نتیجه میتواند خاموش شدن کل سیستم گردش خون باشد.
هنگامی که یک سیستم بدن از کار میافتد، بقیه آنها نیز از کار میافتند زیرا هر سیستمی در بدن شما به نوعی به هم متصل است. همچنین، تنها راهی که ارگانیسمهای چندسلولی ایجاد میشوند، فرایند میتوز است. بدون تقسیم سلولی، انسان و هر موجود چند سلولی دیگر به صورت یک سلول باقی میماند.
انواع تقسیم سلول در موجودات مختلف
تقسیم سلولی فرایندی است که سلولها برای تقسیم شدن طی میکنند. بسته به نوع ارگانیسمی که در حال تقسیم است، انواع مختلفی از تقسیم سلولی وجود دارد. ارگانیسمها در طول زمان تکامل یافتهاند تا اشکال مختلف و پیچیدهتری از تقسیم سلولی داشته باشند. بیشتر پروکاریوتها یا باکتریها از شکافت دوتایی برای تقسیم سلول استفاده میکنند. یوکاریوتها در هر اندازه از میتوز برای تقسیم استفاده میکنند.
یوکاریوتهایی که از نظر جنسی تولید مثل میکنند از شکل خاصی از تقسیم سلولی به نام میوز برای کاهش محتوای ژنتیکی در سلول استفاده میکنند. این در تولید مثل جنسی ضروری است زیرا هر یک از والدین باید فقط نیمی از مواد ژنتیکی مورد نیاز را بدهد، در غیر این صورت فرزندان بیش از حد DNA خواهند داشت که میتواند مشکلساز باشد.
با توجه به این توضیحات میتوان گفت انواع مختلف تقسیم سلولی که در جانداران مختلف وجود دارند، سه مورد زیر هستند.
- شکافت دوتایی: در پروکاریوتها دیده میشود.
- تقسیم میتوز: در یوکاریوتها وجود دارد.
- تقسیم میوز: در یوکاریوتها وجود دارد.

شکافت دوتایی پروکاریوت ها
پروکاریوتها از طریق نوعی تقسیم سلولی به نام شکافت دوتایی تکثیر میشوند. پروکاریوتها ارگانیسمهای سادهای هستند که تنها یک غشای سلولی دارند و در داخل هیچ تقسیم دیگری وجود ندارد. بنابراین، هنگامی که یک موجود پروکاریوت تقسیم میشود، به سادگی DNA را تکثیر میکند و به نصف تقسیم میشود. البته باید گفت که این فرایند کمی پیچیدهتر است، زیرا DNA ابتدا باید توسط پروتئینهای خاصی باز شود.
اگرچه DNA در پروکاریوتها معمولاً در یک حلقه وجود دارد، اما زمانی که سلول از آن استفاده میکند میتواند کاملاً در هم بپیچد. برای کپی شدن موثر DNA، باید مولکول در هم تنیده داخل آن سلول باز شود. این پدیده همچنین اجازه میدهد تا دو حلقه جدید DNA ایجاد شده پس از تولید از هم جدا شوند. دو رشته DNA به دو سمت مختلف سلول پروکاریوت جدا میشوند. سپس سلول کشیدهتر شده و در وسط تقسیم میشود. این فرایند در تصویر زیر قابل مشاهده است.

در این تصویر DNA به صورت خط درهمتنیده است. سایر اجزاء دارای برچسب هستند. پلاسمیدها حلقههای کوچکی از DNA هستند که در طول شکافت دوتایی نیز کپی میشوند و میتوانند در محیط، از سلولهای مرده که از هم جدا میشوند نیز، برداشت شوند. این پلاسمیدها سپس میتوانند بیشتر تکثیر شوند. اگر پلاسمید مفید باشد، در جمعیت افزایش مییابد. این تا حدی نحوه ایجاد مقاومت آنتیبیوتیکی در باکتریها است. ریبوزومها ساختارهای پروتئینی کوچکی هستند که به تولید پروتئین کمک میکنند. آنها همچنین تکثیر میشوند تا هر سلول بتواند به اندازه کافی عملکرد داشته باشد.
تقسیم میتوز یوکاریوت ها
ارگانیسمهای یوکاریوتی دارای اندامکهای متصل به غشاء و DNA هستند که روی کروموزومها وجود دارد و تقسیم سلولی را سختتر میکند. یوکاریوتها باید DNA، اندامکها و مکانیسمهای سلولی خود را قبل از تقسیم تکثیر کنند. بسیاری از اندامکها با استفاده از فرایندی که اساساً شکافت دوتایی است، تقسیم میشوند، که دانشمندان را به این باور رساند که یوکاریوتها در سالهای ابتدایی شکلگیری حیات، توسط پروکاریوتهایی که در داخل پروکاریوتهای دیگر زندگی میکردهاند تشکیل شدهاند.
پس از تکثیر DNA و اندامکها در طول اینترفاز چرخه سلولی، یوکاریوت میتواند فرایند میتوز را آغاز کند. این فرایند در طول پروفاز، زمانی که کروموزومها متراکم میشوند، آغاز میشود. اگر میتوز بدون متراکم شدن کروموزومها پیش میرفت، DNA در هم پیچیده میشد و میشکست. DNA یوکاریوتی با پروتئینهای زیادی مرتبط است که میتواند آن را به ساختارهای پیچیده تبدیل کند.
همانطور که میتوز به سمت متافاز پیش میرود، کروموزومها در وسط سلول قرار میگیرند. هر نیمه از کروموزوم، به کروماتیدهای خواهری معروف است، زیرا کپیهای مشابهی از یکدیگر هستند و با ادامه میتوز در هر نیمه از سلول جدا میشوند. در پایان میتوز، فرایند دیگری به نام سیتوکینزیس سلول را به دو سلول دختر جدید تقسیم میکند. همه موجودات یوکاریوتی از میتوز برای تقسیم سلولهای خود استفاده میکنند. با این حال، تنها موجودات تکسلولی از میتوز به عنوان نوعی تولیدمثل استفاده میکنند.
بیشتر موجودات چندسلولی به روش جنسی در حال تولیدمثل هستند و DNA خود را با موجودات دیگر ترکیب میکنند تا تولیدمثل کنند. در این موارد، موجودات به روش متفاوتی برای انجام تقسیم سلولی نیاز دارند. میتوز سلولهای یکسانی تولید میکند، اما میوز سلولهایی با نیمی از اطلاعات ژنتیکی یک سلول معمولی تولید میکند و به دو سلول از ارگانیسمهای مختلف از یک گونه اجازه میدهد با هم ترکیب شوند.

تقسیم میوز یوکاریوت ها
در جانورانی که از نظر جنسی تولیدمثل میکنند، معمولاً لازم است قبل از لقاح اطلاعات ژنتیکی کاهش یابد. برخی از گیاهان میتوانند با تعداد زیادی کپی از کد ژنتیکی وجود داشته باشند، اما در بیشتر موجودات زنده داشتن نسخههای زیاد بسیار مضر است. انسانهایی که حتی یک نسخه اضافی از یک کروموزوم دارند، میتوانند تغییرات مضری را در بدن خود تجربه کنند. برای مقابله با این مسئله، ارگانیسمهای دارای تولیدمثل جنسی تحت نوعی تقسیم سلولی به نام میوز قرار میگیرند.
مانند قبل از میتوز، DNA و اندامکها همانندسازی میشوند. فرایند میوز شامل دو تقسیم سلولی متفاوت است که پشت سر هم اتفاق میافتند. اولین میوز، یعنی میوز I، کروموزومهای همولوگ را جدا میکند. کروموزومهای همولوگ موجود در یک سلول نشاندهنده دو آلل هر ژنی است که یک موجود زنده دارد. این آللها دوباره ترکیب و جدا میشوند، بنابراین سلولهای دختر حاصل، تنها یک آلل برای هر ژن دارند و هیچ جفت کروموزوم همولوگی ندارند.
بخش دوم، میوز II، دو نسخه از DNA را مانند میتوز جدا میکند. نتیجه نهایی میوز یک سلول، ایجاد ۴ سلول است و هر کدام این این سلولهای جنسی ایجادشده تنها یک نسخه از ژنوم دارند که نصف تعداد طبیعی است. ارگانیسمها معمولاً این سلولها را در گامتها بستهبندی میکنند، که میتوانند برای یافتن گامتهای دیگر به محیط بیرونی سفر کنند.
هنگامی که دو گامت از نوع مناسب به هم میرسند، یکی دیگری را بارور کرده و یک زیگوت تولید میکند. زیگوت یک سلول منفرد است که سپس برای تولید میلیونها سلول لازم برای ایجاد ارگانیسم بزرگ تحت میتوز قرار میگیرد. بنابراین، بیشتر یوکاریوتها از میتوز و میوز اما در مراحل مختلف چرخه زندگی خود استفاده میکنند.

جدول تفاوت های میوز و میتوز
تفاوتهای بین دو نوع تقسیم سلولی میوز و میتوز در جدول زیر خلاصه شده است:
| تقسیم میتوز | تقسیم میوز |
| سلولهای دیپلوئید (2n) تولید میکند. | سلولهای هاپلوئید (n) تولید میکند. |
| شامل یک تقسیم هستهای است. | شامل دو تقسیم هستهای است. |
| محصول آن سلول سوماتیک است. | محصول این نوع تقسیم سلولی گامت است. |
| مسئول تولیدمثل غیرجنسی | مسئول تولیدمثل جنسی است. |
| کراسینگاور وجود ندارد. | کراسینگاور در آن رخ میدهد. |
| دو سلول در آن تولید میشوند. | چهار سلول در آن تولید میشود. |
تقسیم سلولی میتوز و مراحل آن
همانطور که بیان شد میتوز مرحلهای از چرخه سلولی است که در آن کروموزومها در هسته به طور مساوی بین دو سلول تقسیم میشوند. وقتی فرایند تقسیم سلولی کامل شد، دو سلول دختر با مواد ژنتیکی یکسان تولید میشوند. میتوز نحوه تقسیم سلولهای سوماتیک یا غیر تولیدمثلی است. سلولهای سوماتیک بیشتر بافتها و اندامهای بدن شما از جمله پوست، ماهیچهها، ریهها، روده و سلولهای مو را تشکیل میدهند. سلولهای تولیدمثلی (مانند تخمک) سلولهای سوماتیک نیستند.
سلولهای بنیادی مانند روده، فولیکولهای مو و سلولهای بنیادی خونساز، کمترین سرعت را تقسیم سلولی دارند به آرامی تقسیم میشوند تا از جهشهای DNA جلوگیری کرده و سلولهای دختری سالم و تقویت شده بسازند تا در نهایت بتوانند به هومئوستاز بهتر و ترمیم بافت کمک میکنند.
فرایند تقسیم میتوز دارای چندین مرحله یا مراحل از چرخه سلولی برای ساخت موفقیتآمیز سلولهای دیپلوئید جدید است که شامل موارد زیر هستند.
- اینترفاز
- پروفاز
- پرومتافاز
- متافاز
- آنافاز
- تلوفاز
- سیتوکینز
قبل از شروع به تقسیم، سلول مراحل دیگری را نیز طی میکند که در چرخه سلولی بخش «اینترفاز» نامگذاری شده است. در ادامه این مرحله و سایر مراحل تقسیم میتوز را به ترتیب توضیح دادهایم.

اینترفاز و مراحل آن
اینترفاز دورهای است که سلول در حال آماده شدن برای تقسیم و شروع چرخه سلولی است. در این مدت، سلولها در حال جمعآوری مواد مغذی و انرژی هستند. سلول والد همچنین در حال ساخت یک کپی از DNA خود است تا به طور مساوی بین دو سلول دختر تقسیم شود. قبل از اینکه سلول در حال تقسیم وارد میتوز شود، دورهای از رشد به نام اینترفاز را پشت سر میگذارد. حدود ۹۰ درصد از زمان زندگی سلول در چرخه سلولی طبیعی ممکن است در اینترفاز سپری شود. اینترفاز شامل مراحل زیر است.
فاز G1
دوره قبل از سنتز DNA نامگذاری شده است. در این مرحله، سلول برای آماده شدن جهت تقسیم سلولی، جرم خود را افزایش میدهد. فاز G1 اولین وقفه سلولی را در خود دارد. در این مرحله از جنبه میکروسکوپی، تغییر کمی قابل مشاهده است. با این حال، در مرحله G1، سلول در سطح بیوشیمیایی کاملاً فعال است. سلول رشد میکند و بلوکهای ساختمانی DNA کروموزومی و پروتئینهای مرتبط و همچنین ذخایر انرژی کافی را برای تکمیل تکثیر هر کروموزوم در هسته جمعآوری میکند.
فاز S
مرحله سنتز اینترفاز به دلیل پیچیدگی تکثیر ماده ژنتیکی، طولانیترین زمان را میبرد. در طول اینترفاز، DNA هستهای در یک پیکربندی کروماتین نیمهمتراکم باقی میماند. در فاز S، همانندسازی DNA منجر به تشکیل جفتهای یکسان از مولکولهای DNA، کروماتیدهای خواهری، میشود که محکم به ناحیه سانترومری متصل هستند.
سانتروزوم در طول فاز S تکثیر میشود. دو سانتروزوم باعث ایجاد دوک میتوزی میشود و دستگاهی است که حرکت کروموزومها را در طول میتوز هماهنگ میکند. در مرکز هر سلول جانوری، سانتروزوم سلولهای حیوانی با یک جفت اجسام میله مانند به نام سانتریولها که در زوایای قائمه با یکدیگر قرار دارند، در ارتباط هستند. سانتریولها به سازماندهی تقسیم سلولی کمک میکنند. سانتریولها در سانتروزوم سایر گونههای یوکاریوتی مانند گیاهان و بیشتر قارچها وجود ندارند.
فاز G2
دورهای که پس از سنتز DNA رخ داده اما قبل از شروع پروفاز میتوز است. سلول پروتئینها را سنتز میکند و به افزایش اندازه ادامه میدهد. فاز G2 دومین فاز وقفه در چرخه سلولی است. در بخش آخر اینترفاز، سلول هنوز دارای هسته است. هسته توسط یک پوشش هستهای محدود شده است و کروموزومهای سلول تکثیر شده اما به شکل کروماتین هستند. در فاز G2، سلول ذخایر انرژی خود را دوباره پر میکند و پروتئینهای لازم برای دستکاری کروموزوم را سنتز میکند.
برخی از اندامکهای سلولی تکثیر میشوند و اسکلت سلولی از بین میرود تا منابعی برای فاز میتوزی فراهم کند. ممکن است رشد سلولی اضافی در طول G2 وجود داشته باشد. آمادهسازی نهایی برای مرحله میتوز باید قبل از اینکه سلول بتواند وارد مرحله اول میتوز شود به طور کامل انجام گیرد.

پروفاز میتوز
در پروفاز، کروماتین به کروموزومهای مجزا متراکم میشود. پوشش هسته شکسته شده و دوکها در قطبهای مخالف سلول تشکیل میشوند. پروفاز (در مقابل اینترفاز) اولین مرحله واقعی فرایند میتوز است. در طول پروفاز، تعدادی از تغییرات مهم رخ میدهد که شامل موارد زیر هستند:
- الیاف کروماتین به کروموزومها پیچیده میشوند و هر کروموزوم دارای دو کروماتید است که در یک سانترومر به هم متصل میشوند.
- دوک میتوزی متشکل از میکروتوبولها و پروتئینها در سیتوپلاسم تشکیل میشود.
- دو جفت سانتریول (که از تکثیر یک جفت در اینترفاز به وجود میآیند) به دلیل طولانی شدن میکروتوبولهایی که بین آنها تشکیل میشود، از یکدیگر به سمت انتهای مخالف سلول حرکت میکنند.
- الیاف قطبی ریزلولههایی هستند که رشتههای دوک را تشکیل میدهند. این الیاف از هر قطب سلولی به استوای سلول میرسند.
- «کینتوکورها» (Kinetochore) که نواحی تخصصی در سانترومر کروموزومها هستند به نوعی میکروتوبول به نام فیبرهای کینتوکور متصل میشوند.
- الیاف کینتوکورها با الیاف قطبی دوکی که کینتوکورها را به الیاف قطبی متصل میکنند تعامل کرده و کروموزومها شروع به مهاجرت به سمت مرکز سلول میکنند.

متافاز میتوز
در متافاز، دوک به بلوغ میرسد و کروموزومها در صفحه متافازی (صفحهای که به یک اندازه از دو قطب سلول فاصله دارد) تراز میشوند. در متافاز، سانترومرهای کروموزومها روی صفحه متافازی (یا صفحه استوایی) میپیوندند، یک خط فرضی که در فواصل مساوی از دو قطب سانتروزوم قرار دارد و توسط کمپلکسهایی به نام کوهزین در کنار هم قرار میگیرد. در این مرحله، تعدادی از تغییرات رخ میدهد که شامل موارد زیر هستند:
- غشای هستهای به طور کامل ناپدید میشود.
- الیاف قطبی (ریز لولههایی که الیاف دوک را تشکیل میدهند) همچنان از قطبها تا مرکز سلول گسترش مییابند.
- کروموزومها بهطور تصادفی حرکت میکنند تا زمانی که از دو طرف سانترومر خود به فیبرهای قطبی بچسبند (در کینتوکور). کروموزومها در وسط سلول بهوسیله «مراکز سازماندهنده میکروتوبول» (MTOCs) که سانترومرهای هر دو کروماتید را فشار میدهند، کشیده شده و در نتیجه کروموزوم را به سمت مرکز حرکت داده و ردیف میکنند.
- کروموزومها در صفحه متافاز در زوایای قائم با قطبهای دوک قرار دارند. در طول این مرحله، همه میکروتوبولها، به استثنای کینتوکورها، در حالت بیثباتی هستند که باعث پیشرفت آنها به سمت آنافاز میشود.
- کروموزومها در صفحه متافاز توسط نیروهای مساوی الیاف قطبی که به سانترومرهای کروموزومها فشار میآورند نگه داشته میشوند. در این مرحله، کروموزومها آماده تقسیم به قطبهای مخالف سلول به سمت دوک متصل به خود هستند.

آنافاز میتوز
در آنافاز، کروموزومهای جفت شده (کروماتیدهای خواهر) از هم جدا میشوند و شروع به حرکت به سمت انتهای مخالف (قطب) سلول میکنند. الیاف دوکی که به کروماتیدها متصل نیستند سلول را کشیده میکنند. آنافاز مرحله بسیار کوتاهی از چرخه سلولی است و پس از همراستایی کروموزومها در صفحه میتوزی رخ میدهد. کینتوکورها سیگنالهای مهار آنافاز را تا زمان اتصال به دوک میتوزی منتشر میکنند.
هنگامی که کروموزوم نهایی به درستی تراز شد و به آن متصل شد، سیگنال نهایی از بین میرود و باعث تغییر ناگهانی به آنافاز میشود. این تغییر ناگهانی ناشی از فعال شدن کمپلکس ارتقا دهنده آنافاز و عملکرد آن برای برچسب زدن تخریب پروتئینهای مهم در انتقال متافاز-آنافاز است. در پایان آنافاز، هر قطب حاوی مجموعه کاملی از کروموزومها است. در طول آنافاز، تغییرات کلیدی زیر رخ میدهد:
- سانترومرهای جفت شده در هر کروموزوم مجزا شروع به حرکت و دور شدن از هم میکنند.
- هنگامی که کروماتیدهای خواهری جفت شده از یکدیگر جدا میشوند، هر کدام یک کروموزوم کامل در نظر گرفته میشوند. به آنها کروموزوم دختری گفته میشود.
- از طریق دستگاه دوک، کروموزومهای دختری به سمت قطبهای انتهای مخالف سلول حرکت میکنند.
- کروموزومهای دختری ابتدا از سانترومر مهاجرت میکنند و رشتههای کینتوکور مانند کروموزومهای نزدیک به یک قطب کوتاهتر میشوند.
- در آمادهسازی برای تلوفاز، دو قطب سلولی نیز در طول دوره آنافاز از هم دورتر میشوند. در پایان آنافاز، هر قطب حاوی مجموعه کاملی از کروموزومها است.

تلوفاز میتوز
تلوفاز آخرین مرحله از چرخه سلولی است که در آن یک شیار، شکاف سیتوپلاسم سلولی (سیتوکینز) و کروماتین را میشکافد. از طریق سنتز یک پوشش هستهای جدید که در اطراف کروماتین جمع شده در هر قطب تشکیل میشود، رخ میدهد. با بازگشت کروماتین به حالت شلی که در طول فاز میانی داشت، هسته اصلاح میشود. در تلوفاز، کروموزومها به هستههای جدید مجزا در سلولهای دختر در حال ظهور محاصره میشوند.
تغییرات زیر در این مرحله رخ میدهد:
- الیاف قطبی همچنان درازتر میشوند.
- هستهها در قطبهای مخالف شروع به تشکیل میکنند.
- پوششهای هستهای این هستهها از قطعات باقیمانده از پوشش هسته سلول مادر و از قطعات سیستم غشایی داخلی تشکیل میشود.
- هستهها دوباره ظاهر میشوند.
- رشتههای کروماتین کروموزومها باز میشوند.
- پس از این تغییرات، تلوفاز میتوز تا حد زیادی کامل میشود. محتویات ژنتیکی یک سلول به طور مساوی به دو قسمت تقسیم شده است.
برای درک بهتر این مراحل، پیشنهاد میکنیم ویدیوی آموزشی رایگان زیر را مشاهده کنید:

سیتوکینز
سیتوکینز فرایند نهایی در تقسیم سلولی یوکاریوتی است که سیتوپلاسم، اندامکها و غشای سلولی را تقسیم میکند. سیتوکینز معمولاً در انتهای میتوز، پس از تلوفاز رخ میدهد، اما این دو فرایند مستقل هستند. در اکثر حیوانات، سیتوکینز زمانی در اواخر آنافاز یا تلوفاز اولیه شروع میشود تا اطمینان حاصل شود که کروموزومها کاملاً جدا شدهاند. حرکات سیتوکینزیس که در سلول دیده میشود توسط همان شبکه دوکی ایجاد میشود که مسئول جداسازی کروموزومها بود. بخشهایی از دوک که مسئول حرکت کروموزومها هستند در اواخر تقسیم سلولی شکسته میشوند تا در بازسازی دو سلول جدید استفاده شوند.
سلولها میتوانند به طور مساوی تقسیم شوند که به عنوان سیتوکینز متقارن شناخته میشود، یا یکی از سلولها میتواند اکثر سیتوپلاسم را حفظ کند. به عنوان مثال، در طول میوز در جنس نر انسان، هر ۴ سلول در انتهای میوز دارای اندازه و تعداد نسبی اندامک هستند. این فرایند اسپرمزایی میلیونها اسپرم کوچک، اما عمدتاً مساوی تولید میکند. از طرف دیگر اووژنز (تشکیل تخمک) انسانی از طریق سیتوکینز نامتقارن تقسیم میشود. این فرایند یک سلول بسیار بزرگ و ۳ جسم قطبی تولید میکند.
اجسام قطبی کوچکتر به تخم تبدیل نمیشوند. به این ترتیب تخمکهای کمتری تولید میشود اما سلولهای بسیار بزرگتری هستند. برخی از سلولها، در انسان و سایر گونهها، پس از میتوز تحت سیتوکینز قرار نمیگیرند و سلولهای چندهستهای بزرگ را تشکیل میدهند.
سیتوکینز در سلول های جانوری
خواه تقسیم سلولی میتوز باشد یا میوز، سیتوکینز تقریباً به همین صورت اتفاق میافتد. سیگنالهای سلولی به سلول میگویند کجا تقسیم شود، که صفحه تقسیم را ایجاد میکند. در اطراف این صفحه، شیار سیتوکینتیک تشکیل میشود که در نهایت برای جدا کردن دو سلول از هم، کامل شده و سلولها را جدا میکند. فرایند نهایی سیتوکینز در سلولهای جانوری، بریده شدن است. در حین بریدگی، حلقه انقباضی اکتین-میوزین که شیار سیتوکینتیک را ایجاد میکند، در تمام مسیر منقبض میشود و غشاهای پلاسمایی تحت شکافت قرار میگیرند تا در نهایت دو سلول از هم جدا شوند.

دانشمندان هنوز مطمئن نیستند که چه چیزی باعث مشخص شدن صفحه تقسیم در سلولهای مختلف میشود. این یک فرایند پیچیده بوده که شامل بسیاری از میکروتوبولها و سیگنالهای سلولی است. هنگامی که این موقعیت مشخص شد، حلقه انقباضی اکتین-میوزین باید ایجاد شود. اکتین و میوزین همان پروتئینهای حرکتی هستند که باعث انقباض سلولهای عضلانی میشوند. سلولهای عضلانی مملو از رشتههای اکتین هستند که پروتئین میوزین در صورت دریافت انرژی ATP میتواند آنها را جمع کند. همین سیستم در تقسیم سلولهای حیوانی به کار میرود.
نحوه تشکیل حلقه انقباضی در سیتوکینز
رشتههای اکتین یک حلقه در صفحه تقسیم تشکیل میدهند. سپس پروتئینهای میوزین شروع به کشیدن رشتههای اکتین به هم میکنند و حلقه کوچکتری ایجاد میکنند. در نهایت، تمام سیتوپلاسم و اندامکها از حلقه حذف شدند. تنها چیزی که باقی مانده حلقه اکتین-میوزین و میکروتوبولهایی است که توسط حلقه منقبض شدهاند.
این ساختار میانی بدن نامیده میشود و همچنین باید تقسیم شود تا سلولها از هم جدا شوند. این مراحل در طول فرایند «جدایی/بریده شدن» (Abscission) اتفاق میافتد. پروتئینها بریده میشوند و غشای پلاسمایی ذوب میشوند. ماده خارج سلولی که سلولها را در کنار هم نگه میدارد حل میشود و سلولها میتوانند از هم جدا شوند.
در برخی از جانوران چند سلولی، سلولها ارتباط نزدیکی با هم دارند و حتی میتوانند بین سیتوپلاسمهایشان که به نام اتصالات شکافدار شناخته میشوند، ایجاد و حفظ کنند. این پلهای کوچک میتوانند بهعنوان بقایای شبکه آندوپلاسمی که در ساختار میانی بدن به دام میافتند تشکیل شده یا بعداً ساخته شوند.
سیتوکینز در گیاهان چگونه است؟
گیاهان تحت فرایند مشابهی از سیتوکینز قرار میگیرند، با تفاوت در استحکام سلولهای آنها. گیاهان توسط یک لایه ثانویه، دیواره سلولی احاطه شدهاند. این ساختار خارج سلولی مسئول کمک به فرم دادن به گیاهان است و باید هنگام تقسیم سلولی ایجاد شود. برای انجام این کار، گیاهان از ساختارهای دوکی میکروتوبولی به نام فراگموپلاست استفاده میکنند.
فراگموپلاستها وزیکولهایی از مواد دیواره سلولی را به صفحه سلولی جدید حمل میکنند که از دستگاه گلژی آمدهاند. این مواد مانند سلولز برهمکنش میکنند و ماتریکس پیچیده و قوی را تشکیل میدهند. پس از اینکه صفحه سلول را تقسیم کرد، غشای پلاسمایی بسته میشود و دو سلول از هم جدا میشوند.

فراگموپلاست مانند سانتروزوم سلولهای جانوری، میکروتوبولها را سازماندهی میکند و رشد و کاهش آنها را هدایت میکند. اجزای صفحه سلولی جدید توسط شبکه آندوپلاسمی و دستگاه گلژی ایجاد و بستهبندی میشوند. سپس به فراگموپلاست فرستاده میشوند که صفحه سلولی را از وسط به بیرون میسازد. این را میتوان در نمودار بالا مشاهده کرد.
صفحه سلولی از وسط شروع میشود و با تکمیل آن، میکروتوبولهای فراگموپلاست به سمت بیرون حرکت میکنند تا به غشای پلاسمای فعلی برسند. این غشاء بریده میشود و دیواره سلولی به طور کامل بین تمام سلولهای اطراف متصل میشود. بین دو سلول، شبکه آندوپلاسمیک به دام افتاده پلاسمودسماتا ایجاد میکند که مانند اتصالات شکافدار هستند و به مولکولها اجازه میدهند از سلولی به سلول دیگر عبور کنند. این نظریه وجود دارد که گیاهان ممکن است از این پلاسمودسماتا به عنوان شکلی از ارتباطات سلولی استفاده کنند.
تقسیم سلولی میوز و مراحل آن
میتوز تقریباً برای تمام نیازهای تقسیم سلولی بدن شما استفاده میشود. سلولهای جدید را در طول رشد اضافه میکند و سلولهای قدیمی و فرسوده را در طول زندگی موجودات جایگزین میکند. هدف از میتوز تولید سلولهای دختری است که از نظر ژنتیکی مشابه مادران خود هستند و حتی یک کروموزوم کم و بیش ندارند. از سوی دیگر، میوز فقط برای یک هدف در بدن انسان استفاده میشود: تولید گامت یا سلولهای جنسی به نام اسپرم و تخمک. هدف آن ساختن سلولهای دختری با نیمی از کروموزومهای سلول اولیه است.
از بسیاری جهات، میوز بسیار شبیه میتوز است. سلول مراحل مشابهی را طی میکند و از استراتژیهای مشابهی برای سازماندهی و جداسازی کروموزومها استفاده میکند. اما در میوز، سلول وظایف پیچیدهتری دارد. هنوز هم مانند میتوز نیاز به جداسازی کروماتیدهای خواهری (دو نیمه یک کروموزوم) دارد. اما علاوه بر این باید کروموزومهای همولوگ را نیز جدا کند، جفتهای کروموزوم مشابه اما غیریکسانی که یک موجود زنده از دو والدین خود دریافت میکند.
این اهداف در میوز با استفاده از فرایند تقسیم دو مرحلهای انجام میشود. جفتهای همولوگ در طول اولین دور تقسیم سلولی که میوز I نامیده میشود از هم جدا میشوند. کروماتیدهای خواهری در طول دور دوم که میوز II نامیده میشود، جدا میشوند. از آنجایی که تقسیم سلولی دو بار در طول میوز اتفاق میافتد، یک سلول شروعکننده میتواند چهار گامت (تخمک یا اسپرم) تولید کند. در هر دور تقسیم، سلولها چهار مرحله را طی میکنند.
- پروفاز
- متافاز
- آنافاز
- تلوفاز
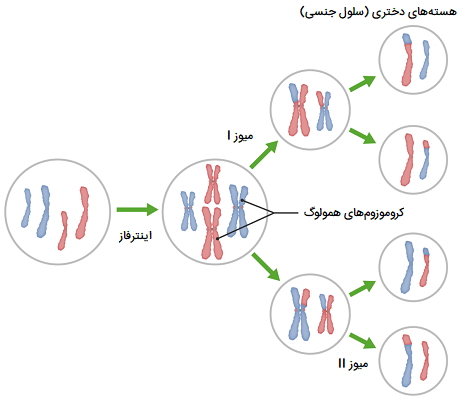
در این تصویر تقسیم میوز و مراحل آن نشان داده شده است.

مراحل میوز I چیست؟
در چرخه سلولی، میوز I پس از اینترفاز که کروموزومها در فاز S تکثیر میشوند، اتفاق میافتد. سپس، کروموزومها در مراحل اولیه پروفاز I متراکم میشوند. دو سانتروزوم به دو قطب مخالف سلول میروند و آن را برای تقسیم هستهای آماده میکنند. کروموزومهای همولوگ از جفت کروماتید تشکیل شدهاند. این کروموزومها پس از جفت شدن بیوالانت (تتراد) تشکیل میدهند تا در طول متافاز I در خط استوای دوک قرار گیرند. اگرچه کروموزومهای همولوگ در طول آنافاز I از یکدیگر جدا میشوند، اما دو کروماتید خواهری به هم متصل میمانند.

پروفاز میوز I
پروفاز I پیچیدهترین مرحله میوز I است که بیشتر به پنج مرحله تقسیم میشود که عبارتند از: لپتوتن، زیگوتن، پاکیتن، دیپلوتن و دیاکینزیس. در ادامه هر کدام از این مراحل را توضیح دادهایم.
- مرحله لپتوتن با متراکمشدن الیاف کروماتین شروع میشود که شبیه ساختار تشکیلشده در ابتدای میتوز است.
- مرحله زیگوتن شامل تراکم بیشتر الیاف است که آنها را قادر میسازد به عنوان کروموزومهای منفرد متمایز شوند. در نتیجه سیناپسیس (قرارگیری کروموزومهای همولوگ هنگام میوز)، رخ میدهد و تترادها زمانی تشکیل میشوند که جفت کروموزومها با هم جفت شوند.
- در مرحله پاکیتن تشکیل تترادها صورت میگیرد که در فرایند تبادل بخشهای DNA حاوی ماده ژنتیکی بین دو کروموزوم نزدیک در فرایندی به نام کراسینگاور بسیار مهم است. بخشهای مربوط به کروموزومها اطلاعات ژنتیکی را برای ترکیب مجدد ژنها (نوترکیبی) مبادله میکنند. متراکم شدن کروموزومها تا تقریباً کمتر از یک چهارم طول آن در مرحله پاکیتن نیز اتفاق میافتد.
- در مرحله دیپلوتن، در نزدیکی سانتروزوم، دو کروموزوم هر تتراد از یکدیگر جدا میشوند. با این حال، دو کروموزوم توسط کیاسماها متصل میمانند، این دو کروموزوم در محلی هستند که در آن دو کروموزوم همولوگ بخشهای DNA را مبادله میکنند.
در طی دیپلوتن، رونویسی از سر گرفته میشود، کروموزومها متراکم میشوند و سلول میوز را برای مدت معینی متوقف میکند. در آغاز مرحله نهایی پروفاز I، دیاکینزیس، زمانی که کروموزومها مجدداً متراکم میشوند تا حداکثر حالت تراکم خود را داشته باشند، سانتروزومها بیشتر حرکت میکنند. کروموزومها فقط توسط کیاسماتا متصل میشوند. در اینجا دوکها تشکیل میشوند، هستهها ناپدید شده و پوشش هستهای از بین میرود. شروع تشکیل دوک میوز و متلاشی شدن هستهها نشانه پایان پروفاز میوز I و شروع متافاز میوز I است.

متافاز میوز I
در طی این مرحله، تترادها پس از اتصال به میکروتوبولها با استفاده از کینتوکور خود به استوای دوک حرکت میکنند. این فرایند حرکت تتراد به سمت استوای سلول معمولاً فقط به میوز I محدود میشود و در تقسیم میتوزی رخ نمیدهد. در هر بیوالانت چهار کروماتید وجود دارد، در نتیجه هر تتراد شامل چهار کینتوکور نیز است. این کینتوکورها نزدیک به یکدیگر ظاهر شده و به صورت یک واحد منفرد رو به قطب یکسان سلول ظاهر میشوند.
چنین آرایشی امکان اتصال هر کینتوکور را به میکروتوبولهای قطب دوک در طرف مقابل میدهد. این ترتیب اولین مرحلهای است که برای جداسازی کروموزومها در طول آنافاز بعدی تنظیم میشود. در این مرحله، تترادها به طور تصادفی مرتب میشوند، بر این اساس، کروموزومهای پدری و مادری در یک قطب سلول قرار میگیرند و بنابراین، هر سلول دختر تازه تشکیلشده مخلوطی از کروموزومهای پدری و مادری را در طول حرکت خود به قطبهای مخالف در طول آنافاز دریافت میکند.

آنافاز میوز I
اولین مرحله در آنافاز شامل مهاجرت کروموزومهای همولوگ به قطبهای دوک با کمک کینتوکور آنها است. این مرحله نشاندهنده یکی از تفاوتهای اصلی بین میوز و میتوز است. در طول میتوز، کروماتیدهای خواهری از هم جدا شده زیرا به قطبهای مخالف کشیده میشوند. در میوز I، دو کروماتید خواهری به هم چسبیده میمانند و کروموزومهای همولوگ پس از جدا شدن به سمت قطبهای دوک حرکت میکنند. این مرحله منجر به حضور تعداد هاپلوئید کروموزوم در هر قطب دوک در انتهای آنافاز میوز I میشود.
فرایند جداسازی کروماتید در طول میتوز با جدا کردن دو کروماتید خواهری با کمک یک آنزیم فعالشده به نام «سپاراز» (Separase) انجام میشود. برای متوقفکردن عمل جداسازی در میوز، سلول پروتئین خاصی به نام «شوگوشین» (Shugoshin) تولید میکند که با محافظت از محل سانتروزومی کروموزوم که در آن فرایند برش اتفاق میافتد، از جدا شدن کروماتیدها جلوگیری میکند.
تلوفاز میوز I
فاز نهایی میوز I تلوفاز I است که با مهاجرت کروموزومها به قطبهای دوک مشخص میشود. یک پوشش هستهای میتواند در اطراف کروموزومها قبل از سیتوکینز تشکیل شود تا دو سلول دختر از مجموعه کروموزوم هاپلوئید تولید کند. بیشتر اوقات، کروموزومها پس از شروع میوز II متراکم میشوند. در پایان میوز I، سیتوکینزیس به تولید دو سلول که هر کدام دارای یک هسته هاپلوئید هستند منجر میشود. کروموزومهای هر سلول هاپلوئید هر کدام از دو کروماتید متصل به سانترومر تشکیل شده است.

مراحل میوز II
اینترفاز میوز بعد از پایان میوز I و قبل از شروع میوز II شروع میشود. این مرحله با تکثیر DNA مرتبط نیست زیرا هر کروموزوم قبلاً از دو کروماتید تشکیل شده است که قبل از شروع میوز I توسط فرایند سنتز DNA همانندسازی شدهاند. به طور خلاصه، DNA قبل از شروع میوز I فقط در یک زمان تکثیر میشود. مرحله میوز II یا تقسیم دوم میوزی هدفی مشابه با میتوز دارد که در آن دو کروماتید جدید در دو سلول دختر جدید جهتگیری میشوند. بنابراین، تقسیم میوز دوم را گاهی اوقات به عنوان تقسیم میوز جداسازی نیز میگویند. در ادامه مراحل میوز II را به ترتیب توضیح دادهایم.
پروفاز میوز II
پروفاز II مرحلهای است که به دنبال میوز I یا اینترکینزیس میآید. با ضخیم شدن و کوتاه شدن کروماتیدها در پروفاز II، پوشش هسته و فروپاشی هسته مشخص میشود و سانتروزومها تکثیر شده و به سمت قطبی مهاجرت میکنند. پروفاز دوم سادهتر و کوتاهتر از پروفاز I بوده و به نوعی شبیه پروفاز میتوزی است. از طرف دیگر، پروفاز II با پروفاز I متفاوت است زیرا عبور کروموزومها فقط در طول پروفاز I اتفاق میافتد و نه پروفاز II. متافاز دوم در پایان مرحله پروفاز II شروع میشود.
متافاز II
متافاز II تقسیم میوز نیز مشابه متافاز تقسیم میتوزی است، با این حال، تنها نیمی از تعداد کروموزومها در متافاز II وجود دارد، متافاز II با همترازی کروموزومی در مرکز سلول مشخص میشود.
آنافاز II
مرحلهای است که بعد از متافاز II میآید، در این مرحله کروماتیدهای خواهری جدا شده و به سمت قطبهای سلول حرکت میکنند. آنافاز II شبیه آنافاز میتوزی است که هر دو شامل جداسازی کروماتیدها هستند. کوتاه شدن کینتوکور منجر به حرکت کروماتیدهای خواهری به دو انتهای سلول میشود.

تلوفاز II
تلوفاز آخرین مرحله میوز است، در طول تلوفاز II، چهار سلول هاپلوئید از دو سلول تولید شده در طی میوز I تولید میشوند، غشای هستهای سلولهای تازه تشکیل شده کاملاً توسعه یافته و سلولها در پایان این مرحله کاملاً از هم جدا میشوند. با این حال، در طول اسپرمزایی در انسان و سایر حیوانات، اسپرمها در انتهای تلوفاز II به طور کامل عمل نمیکنند، زیرا برای عملکرد مناسب نیاز به رشد تاژک دارند.
چهار سلول هاپلوئید پس از تلوفاز II و سیتوکینز تولید میشوند، هر سلول دختر دارای تنها یک کروموزوم از دو جفت همولوگ است. سلولهای هاپلوئید تولیدشده حاوی ترکیبی از اطلاعات ژنتیکی کروموزومهای مادر و پدر است. این سلولها به تنوع ژنتیکی در بین افراد یک گونه و همچنین روند تکاملی موجودات کمک میکنند.
تقسیم سلولی میوز در گیاهان و جلبک ها
گیاهان و جلبکها ارگانیسمهای چندسلولی هستند که در چرخه زندگی خود هر دو شکل هاپلوئید و دیپلوئید سلولها را نشان میدهند. این پدیده را تناوب نسلها مینامند که در آن هاگ هاپلوئید توسط میوز تولید میشود. همچنین به همین دلیل است که در گیاهان و جلبکها به آن میوز اسپوریک میگویند. هاگهای تشکیلشده، جوانه میزنند و تحت تقسیم میتوزی قرار میگیرند و باعث ایجاد یک گیاه هاپلوئید یا جلبک هاپلوئید میشوند.
گامتها با تقسیم میتوزی از سلولهای هاپلوئید موجود تولید میشوند. بنابراین، شکل هاپلوئید گامتوفیت نامیده میشود. گامتها در طول لقاح به هم میپیوندند و شکل دیپلوئیدی سلولها را تولید میکنند. هاگها از فرم دیپلوئیدی توسط میوز تشکیل میشوند. بنابراین فرم دیپلوئید را اسپوروفیت مینامند.

تقسیم سلولی میوز در قارچ ها
قارچها همچنین دارای مراحل غیرجنسی و جنسی در چرخه زندگی خود هستند. میسلیوم، بهویژه، ممکن است وارد فاز جنسی یا غیرجنسی شود. هنگامی که میسلیوم هاپلوئید وارد فاز جنسی میشود، تحت پلاسموگامی (همجوشی دو پروتوپلاست) و کاریوگامی (همجوشی دو هسته هاپلوئید) قرار میگیرد. بنابراین، به دنبال کاریوگامی، زیگوت دیپلوئید تشکیل میشود.
زیگوت به صورت اسپورانژیوم ساقهدار رشد میکند که سپس از طریق میوز هاگهای هاپلوئید تشکیل میدهد. اسپورهای تولیدشده توسط میوز در مقابل میتوسپورهایی که از طریق میتوز تولید میشوند، میوسپور نامیده میشوند. این هاگهای هاپلوئید (سلولهای تولیدمثلی) از اسپورانژیوم آزاد میشوند و هر کدام در نهایت به یک میسلیوم جدید جوانه میزنند. بنابراین، در قارچها، میوز سومین مرحله در مراحل متوالی مرحله جنسی است که در آن پلاسموگامی اولین مرحله است و پس از آن کاریوگامی است. میوز در بازگرداندن حالت هاپلوئید قارچ بسیار مهم است.
میوز در انسان و سایر جانوران
میوز گامتهای هاپلوئید را در انسان و سایر حیوانات تولید میکند که بخش مهمی از گامتوژنز است. همانطور که از نام آن پیداست، گامتوژنز فرایند بیولوژیکی ایجاد گامت است. در انسان و سایر حیوانات، دو شکل گامتوژنز وجود دارد.
- اسپرماتوژنز: تشکیل گامت نر، یعنی سلول اسپرم
- اووژنز: تشکیل گامت ماده، یعنی تخمک یا سلول تخمک
در اووژنز، چهار سلول گامت هاپلوئید از یک سلول دیپلوئید تولید میشود. با این حال، تنها یک سلول زنده میماند و به عنوان یک تخمک عمل میکند. سه تای دیگر به اجسام قطبی تبدیل میشوند. این اثر ناشی از تقسیم نابرابر تخمک توسط میوز است که در آن یکی از سلولهای تشکیلشده بیشتر سیتوپلاسم سلول مادر را دریافت میکند، در حالی که سایر سلولهای تشکیلشده تخریب میشوند که به افزایش غلظت مواد مغذی در تخم کمک میکند.

در اسپرماتوژنز، اسپرم ویژگیهای تخصصی خود را به دست میآورد تا پس از میوز و رویدادهای پس از میوز به یک گامت عملکردی تبدیل شود. اسپرمزایی که در آن سلول اسپرم با به دست آوردن یک تاژک عملکردی و از بین بردن بیشتر سیتوپلاسم خود برای تشکیل یک سر فشرده بالغ میشود.
میوز در مرحله تولیدمثل ارگانیسم رخ میدهد اما، در انسان، تقسیم میوز در مراحل مختلفی انجام میگیرد. به عنوان مثال، در مردان، از دوران بلوغ شروع میشود و در طول زندگی آنها ادامه مییابد. در دختران، نوزاد تازه متولد شده تخمکهای اولیه را در پروفاز I متوقف میکند و مراحل بعدی میوز را در دوران بلوغ ادامه میدهد. با این حال، همانطور که هر تخمک اولیه در زمان تخمکگذاری به یک تخمک ثانویه تبدیل میشود، دوباره در متافاز میوز II متوقف میشود.
میوز در زنان تنها در زمان لقاح ادامه مییابد و کامل میشود. اگر تخمک بارور نشود، میوز دیگر ادامه نخواهد داشت و تخمک ثانویه متوقفشده متلاشی میشود و همراه با بافت دیواره رحم در طی قاعدگی بیرون رانده شده و از بین میرود.
خطاها در تقسیم سلولی میوز
میوز مستعد خطا است و بنابراین میتواند بر توانایی انسان در تولیدمثل تأثیر بگذارد. میوز غیرطبیعی تأثیر منفی زیادی بر بقای نسل بشر دارد. خطا در مراحل میوز ممکن است منجر به ناباروری و همچنین تشکیل گامتهایی با ویژگیهای نامتعادل ژنتیکی شود. خطاهای میوز عامل اصلی ناهنجاریهای مادرزادی ناشی از اختلالات ژنتیکی و همچنین ناهنجاریهای ذهنی کودکان تازهمتولد شده هستند.
خطاها در جفت شدن و نوترکیب شدن کروموزومها در بیش از ۳۰ درصد از پاکیتنهای تخمک انسانی وجود دارد که در آن جفتشدن کروموزومهای همولوگ در پدیدهای به نام «آسیناپسیس» (Asynapsis) با شکست مواجه میشود. در مخمر، شکست در جفت شدن کروموزومی میتواند منجر به مرگ سلولی پس از تحریک نقاط بازرسی سلول شود. همین پدیده در سلولهای زایشی انسان نیز مشاهده میشود. در نتیجه، افزایش تخمکها با خطا در جفت شدن کروموزومی منجر به کاهش تعداد سلولهای زاینده شده که منجر به یائسگی زودرس در زنان میشود.

به همین ترتیب، خطا در مراحل میوز تولید اسپرماتوسیت به دلیل کاهش تعداد اسپرمهای عملکردی تولید شده، منجر به ناباروری در مردان میشود. کاهش تعداد سلولهای زاینده در زنان بیشتر از مردان است زیرا مرد روزانه حدود ۳۰۰ تا ۴۰۰ میلیون اسپرم تولید میکند در حالی که زنان در طول زندگی خود حدود ۳۰۰ تا ۴۰۰ عدد تخمک تولید میکنند.
در متافاز I، جفتهای کروموزوم ممکن است به درستی از هم عبور نکنند، بنابراین کروموزومهای جفتنشده بهطور تصادفی با افزایش خطر تولید گامت آنیوپلوئید جدا میشوند، که شامل تعداد نامتعادل کپی کروموزوم است. علاوه بر این، اسپرماتوسیتها ممکن است با آپوپتوز یا نکروز به دلیل عبور ناموفق از بین بروند.
نوترکیبی در تقسیم میوز
گامتهای تولید شده در میوز همگی هاپلوئید هستند، اما از نظر ژنتیکی یکسان نیستند. به عنوان مثال محصولات میوز را برای یک سلول با 2n = 4n، در نظر بگیرید. هر گامت یک نمونه منحصربهفرد از ماده ژنتیکی حاضر در سلول را دارد. در واقع انواع گامت بالقوه بسیار بیشتری نسبت به چهار مورد نشان داده شده در یک نمودار مراحل میوز وجود دارد، حتی برای سلولی که تنها چهار کروموزوم دارد. دو دلیل اصلی که ما میتوانیم گامتهای ژنتیکی متفاوتی داشته باشیم عبارتند از:
- کراسینگ اور: نقاطی که همولوگها روی آنها عبور میکنند و مواد ژنتیکی را مبادله میکنند، کم و بیش بهطور تصادفی انتخاب میشوند و در هر سلولی که از طریق میوز عبور میکند، متفاوت خواهند بود.
- جهتگیری تصادفی جفتهای همولوگ: جهتگیری تصادفی جفتهای همولوگ در متافاز I امکان تولید گامتهایی با مجموعههای مختلف کروموزومهای همولوگ را فراهم میکند.
در یک سلول انسانی، جهتگیری تصادفی جفتهای همولوگ به تنهایی امکان بیش از ۸ میلیون گامت مختلف را فراهم میکند. هنگامی که کراسینگاور را نیز با این تنوع همراه کنیم، تعداد گامتهای ژنتیکی متفاوتی که شما یا هر شخص دیگری میتوانید بسازید، بینهایت خواهد بود.

مراحل شکافت دوتایی در پروکاریوت ها
پروکاریوتها مانند باکتریها از طریق «شکافت دوتایی» (Binary Fission) تکثیر میشوند. برای موجودات تکسلولی، تقسیم سلولی تنها روشی است که برای تولید افراد جدید استفاده میشود. در هر دو سلول پروکاریوتی و یوکاریوتی، نتیجه تولیدمثل سلولی یک جفت سلول دختر است که از نظر ژنتیکی با سلول مادر یکسان هستند. در موجودات تکسلولی، سلولهای دختری افراد حقیقی گونه هستند.
به دلیل سادگی نسبی پروکاریوتها، فرایند تقسیم سلولی یا شکافت دوتایی، فرایندی کمتر پیچیده و بسیار سریعتر از تقسیم سلولی در یوکاریوتها است. کروموزوم DNA منفرد و دایرهای باکتری در هسته محصور نشده است، اما در عوض مکان خاصی به نام نوکلوئید را در داخل سلول اشغال میکند. اگرچه DNA نوکلوئید با پروتئینهایی مرتبط است که به بستهبندی مولکول در اندازه فشرده کمک میکند.
هیچ پروتئین هیستونی و بنابراین، هیچ نوکلئوزومی در پروکاریوتها وجود ندارد. با این حال، پروتئینهای بستهبندی کننده در باکتریها مربوط به پروتئینهای کوهزین و کندانسین هستند که در فشردگی کروموزوم یوکاریوتها نقش دارند. در ادامه مراحل شکافت دوتایی باکتریها را به ترتیب بیان کردهایم.
- تکثیر کروموزوم دایرهای باکتریایی در محل آغاز همانندسازی DNA از هر دو جهت صورت میگیرد. با تشکیل رشتههای دوتایی جدید، هر نقطه مبدا از اتصال دیواره سلولی به سمت انتهای مخالف سلول دور میشود. با افزایش طول سلول، غشای در حال رشد به انتقال کروموزوم ها کمک می کند.
- سلول شروع به طویل شدن میکند و پروتئینهای FtsZ به سمت ناحیه میانی در داخل سلول مهاجرت میکنند. با افزایش طول سلول، غشای در حال رشد به انتقال کروموزوم ها کمک میکند.
- کروموزومهای تکثیر شده شروع به جداسازی و دور شدن از هم میکنند و به انتهای قطبین سلول کشیده شده میروند. پس از اینکه کروموزومها از نقطه میانی سلول کشیده حرکت کردند، جداسازی سیتوپلاسمی آغاز میشود. پروتئینهای FtsZ یک دایره را در اطراف ناحیه میانی سلول بین کروموزومها تشکیل میدهند. تشکیل یک حلقه متشکل از واحدهای تکراری یک پروتئین، FtsZ، تقسیم بین نوکلوئیدها را هدایت میکند.
- دایره ایجاد شده توسط FtsZ تشکیل تیغه تقسیمکننده سلولی را هدایت میکند. مواد مربوط به غشای پلاسمایی و دیواره سلولی تجمع مییابند. تشکیل حلقه FtsZ باعث تجمع پروتئینهای دیگری میشود که با هم کار میکنند تا مواد غشایی و دیواره سلولی جدید را به محل جذب کنند.
- یک سپتوم بین نوکلوئیدها تشکیل میشود که به تدریج از محیط به سمت مرکز سلول امتداد مییابد. هنگامی که دیوارههای سلولی جدید در جای خود قرار میگیرند، سلولهای دختر از هم جدا میشوند. FtsZ در سیتوپلاسم سلولهای جدید ناپدید میشود.
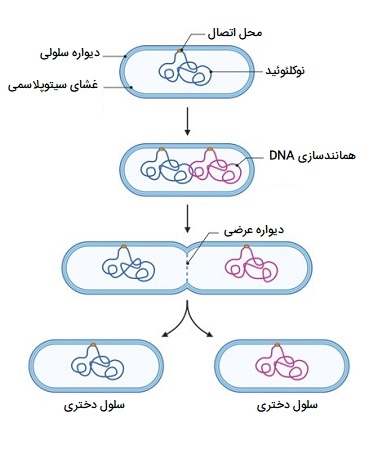
در این تصویر تقسیم دوتایی یا شکافت دوتایی که باعث تکثیر باکتریها میشود نشان داده شده است.

مقایسه شکافت دوتایی و میتوز
شکافت دوتایی باکتری از برخی جهات شبیه میتوز است که در انسان و سایر یوکاریوتها اتفاق میافتد. در هر دو مورد، کروموزومها کپی و جدا میشوند و سلول سیتوپلاسم خود را برای تشکیل دو سلول جدید تقسیم میکند. با این حال، مکانیک و ترتیب این دو فرایند نسبتاً متفاوت است. هیچ دوک میتوزی در باکتریها تشکیل نمیشود. شاید مهمتر از آن، همانندسازی DNA در واقع همزمان با جداسازی DNA در طی شکافت دوتایی اتفاق میافتد (بر خلاف میتوز، جایی که DNA در فاز S، مدتها قبل از جدا شدن در فاز M کپی میشود).
کنترل تقسیم سلولی چگونه است؟
اشتباه در تکثیر یا توزیع کروموزومها منجر به جهشهایی میشود که ممکن است به هر سلول جدید تولید شده از سلول غیرطبیعی منتقل شود. برای جلوگیری از ادامه تقسیم یک سلول در معرض خطر، مکانیسمهای کنترل داخلی وجود دارد که در سه نقطه بازرسی چرخه سلولی اصلی عمل میکنند که چرخه سلولی را میتوان تا زمانی که شرایط مساعد شود متوقف کرد.
- اولین نقطه بازرسی (G1) تعیین میکند که آیا همه شرایط برای ادامه تقسیم سلولی مساعد هستند یا خیر. این نقطه بازرسی نقطهای است که سلول به طور غیرقابل برگشتی به فرایند تقسیم سلولی متعهد میشود. علاوه بر ذخایر کافی و اندازه سلول، بررسی آسیب به DNA ژنومی نیز وجود دارد. سلولی که تمام الزامات را برآورده نکند به فاز S رها نمیشود.
- نقطه بازرسی دوم (G2) ورود به فاز میتوزی را در صورت عدم رعایت شرایط خاص مسدود میکند. مهمترین نقش این ایست بازرسی این است که اطمینان حاصل شود که همه کروموزومها همانندسازی شدهاند و DNA همانندسازی شده آسیبی نمیبیند.
- نقطه بازرسی نهایی (M) در وسط میتوز رخ میدهد. این نقطه بازرسی تعیین میکند که آیا همه کروموزومهای کپیشده بهطور مناسبی چیده شدهاند تا در دو طرف سلول جدا شوند. اگر این به درستی اتفاق نیفتد، میتوان تعداد نادرستی از کروموزومها را در هر یک از سلولهای دختر تقسیم کرد که احتمالاً باعث مرگ آنها میشود.
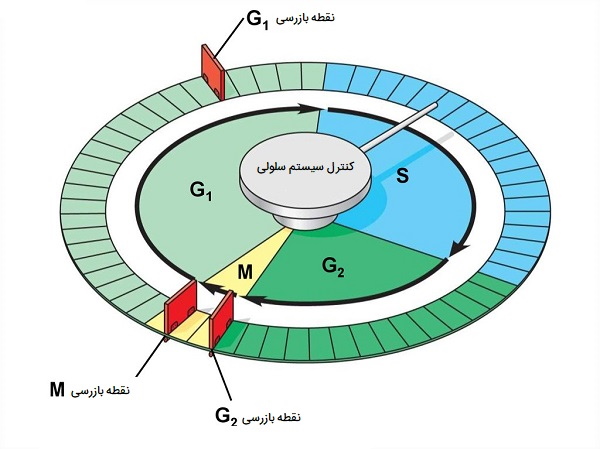
در این تصویر نمایی شماتیک از چرخه سلولی و نقاط بازرسی چرخه رشد و تقسیم سلولی نشان داده شده است.

مولکول های تنظیم کننده چرخه سلولی
علاوه بر نقاط کنترل داخلی، دو گروه از مولکولهای درون سلولی وجود دارند که چرخه سلولی را تنظیم میکنند. این مولکولهای تنظیمکننده یا پیشرفت سلول را به مرحله بعدی فرستاده (تنظیم مثبت) یا چرخه سلولی را متوقف میکنند (تنظیم منفی). مولکولهای تنظیمکننده ممکن است به صورت جداگانه عمل کنند، یا میتوانند بر فعالیت یا تولید سایر پروتئینهای تنظیمکننده تأثیر بگذارند.
بنابراین، ممکن است شکست یک تنظیمکننده واحد تقریباً هیچ تأثیری بر چرخه سلولی نداشته باشد، به خصوص اگر بیش از یک مکانیسم رویداد خاصی را کنترل کنند. همچنین ممکن است در صورت تحت تاثیر قرار گرفتن چندین فرایند، اثر یک تنظیمکننده ناقص یا ناکارآمد بتواند دامنه وسیعی داشته باشد و احتمالاً برای سلول کشنده باشد. در ادامه هر دو نوع تنظیم مثبت و منفی را بررسی کردهایم.

تنظیم مثبت چرخه سلولی
دو گروه از پروتئینها به نامهای سایکلین و کینازهای وابسته به سایکلین (Cdks) مسئول پیشرفت سلول از طریق نقاط بازرسی مختلف هستند. سطوح چهار پروتئین سایکلین در طول چرخه سلولی در یک الگوی قابل پیشبینی در نوسان است. افزایش غلظت پروتئینهای سایکلین توسط سیگنالهای خارجی و داخلی ایجاد میشود. پس از انتقال سلول به مرحله بعدی چرخه سلولی، سایکلینهایی که در مرحله قبل فعال بودند، تجزیه میشوند.
سایکلینها چرخه سلولی را تنها زمانی تنظیم میکنند که محکم به Cdks متصل باشند. برای فعال بودن کامل، کمپلکس Cdk/cyclin نیز باید در مکانهای خاص فسفریله شود. مانند همه کینازها، Cdks آنزیمهایی (کینازها) هستند که سایر پروتئینها را فسفریله میکنند. فسفوریلاسیون پروتئین را با تغییر شکل آن فعال میکند.
پروتئینهای فسفریله شده توسط Cdks در پیشبرد سلول به فاز بعدی نقش دارند. سطح پروتئین Cdk در طول چرخه سلولی نسبتاً پایدار است. با این حال، غلظت سایکلین در نوسان است و تعیین میکند که مجتمعهای Cdk/cyclin چه زمانی تشکیل میشوند. سایکلینها و Cdksهای مختلف در نقاط خاصی در چرخه سلولی متصل میشوند و بنابراین نقاط بازرسی مختلف را تنظیم میکنند.
از آنجایی که نوسانات چرخهای سطوح سایکلین بر اساس زمان چرخه سلولی است و نه بر اساس رویدادهای خاص، تنظیم چرخه سلولی معمولاً توسط مولکولهای Cdk به تنهایی یا مجتمعهای Cdk/cyclin انجام میشود. بدون غلظت خاصی از کمپلکسهای cyclin/Cdk کاملا فعالشده، چرخه سلولی نمیتواند از طریق نقاط بازرسی ادامه یابد.
تنظیم منفی چرخه سلولی
دسته دوم مولکولهای تنظیمکننده چرخه سلولی، تنظیمکنندههای منفی هستند. در تنظیم مثبت، مولکولهای فعال مانند کمپلکسهای CDK/cyclin باعث پیشرفت چرخه سلولی میشوند. در تنظیم منفی، مولکولهای فعال چرخه سلولی را متوقف میکنند. بهترین مولکولهای تنظیمکننده منفی شناختهشده پروتئین رتینوبلاستوما (Rb)، p53 و p21 هستند. بسیاری از آنچه در مورد تنظیم چرخه سلولی شناخته شده است از تحقیقات انجامشده بر روی سلولهایی است که کنترل تنظیمی خود را از دست دادهاند یافته شده است.

هر سه این پروتئینهای تنظیمی در سلولهایی که شروع به تکثیر غیرقابل کنترل کرده بودند (سرطانی شده بودند) به صورت آسیبدیده یا غیرعملکردی کشف شدند. در هر مورد، علت اصلی پیشرفت کنترلنشده در چرخه سلولی، کپی معیوب پروتئین تنظیمکننده بود. به همین دلیل، Rb و سایر پروتئینهایی که به طور منفی چرخه سلولی را تنظیم میکنند، گاهی اوقات سرکوبکننده تومور نامیده میشوند. Rb، p53، و p21 در درجه اول در ایست بازرسی G1 عمل میکنند. در ادامه نحوه عملکرد هر کدام را بیشتر بررسی کردهایم.
عملکرد p53 در چرخه سلول چگونه است؟
p53 یک پروتئین چند عملکردی است که تأثیر عمدهای بر تعهد یک سلول به تقسیم دارد، زیرا این پروتئین زمانی عمل میکند که DNA آسیبدیده در سلولهایی که تحت فرایندهای آمادهسازی در طول G1 هستند. اگر DNA آسیبدیده شناسایی شود، p53 چرخه سلولی را متوقف میکند و آنزیمهایی را برای ترمیم DNA به کار میگیرد. اگر DNA قابل ترمیم نباشد، p53 میتواند باعث آپوپتوز یا خودکشی سلولی شود تا از تکثیر کروموزومهای آسیبدیده جلوگیری شود.
عملکرد p21 در چرخه سلولی
با افزایش سطح p53، تولید p21 آغاز میشود. p21 با اتصال به کمپلکسهای Cdk/cyclin و مهار آن، چرخه دیکتهشده توسط p53 را متوقف میکند. همانطور که یک سلول در معرض استرس بیشتری قرار میگیرد، سطوح بالاتری از p53 و p21 تجمع مییابد و احتمال اینکه سلول به فاز S حرکت کند کمتر میشود.

نحوه عملکرد Rb در چرخه سلولی
Rb تأثیر تنظیمی خود را بر سایر پروتئینهای تنظیمکننده مثبت اعمال میکند. به طور عمده، Rb اندازه سلول را کنترل میکند. در حالت فعال و دفسفریله شده، Rb به پروتئینهایی به نام فاکتورهای رونویسی متصل میشود. فاکتورهای رونویسی ژنهای خاص را «روشن» میکنند و امکان تولید پروتئینهای کدگذاری شده توسط آن ژن را فراهم میکنند. هنگامی که Rb به فاکتورهای رونویسی متصل میشود، تولید پروتئینهای لازم برای انتقال G1/S مسدود میشود.
با افزایش اندازه سلول، Rb به آرامی فسفریله میشود تا زمانی که غیرفعال شود. Rb فاکتورهای رونویسی را آزاد میکند، که اکنون میتواند ژن تولیدکننده پروتئین انتقالی را فعال کند و این بلوک خاص حذف میشود. برای اینکه سلول از کنار هر یک از ایستهای بازرسی عبور کند، همه تنظیمکنندههای مثبت باید «روشن» و همه تنظیمکنندههای منفی «خاموش» باشند.

توده های سرطانی حاصل کدام نوع تقسیم سلولی هستند ؟
به طور معمول، سرطان یک بیماری مربوط به میتوز است. در این حالت، نقاط بازرسی طبیعی که میتوز را تنظیم میکنند توسط سلولهای سرطانی نادیده گرفته میشوند. این شرایط در رویدادی شروع میشود که در آن یک سلول منفرد از یک سلول طبیعی به یک سلول سرطانی (در نتیجه تغییرات در عملکرد یکی از ژنهایی که در کنترل رشد نقش دارند)، تبدیل میشود.
سلولهای سرطانی تمایل دارند که مهار وابسته به تراکم رشد طبیعی را نادیده بگیرند. آنها پس از تماس با سلولهای دیگر تکثیر میشوند و تا زمانی که تمام مواد مغذی تمام شوند به انباشته شدن ادامه میدهند. وقتی صحبت از تقسیم سلولی به میان میآید، این سلولها تقریباً همه قوانین را زیر پا میگذارند. در ادامه ویژگیهای تقسیم سلولهای سرطانی را بررسی میکنیم.
- سلولهای سرطانی میتوانند بدون سیگنالهای خارجی مناسب تقسیم شوند. این مشابه حرکت خودرویی است که بدون اعمال فشار بر پدال گاز حرکت میکند. یک مثال میتواند رشد سلول سرطان سینه بدون نیاز به استروژن (فاکتور رشد طبیعی) باشد. برخی از سلولهای سرطان سینه در واقع با خاموش کردن بیان گیرنده استروژن در سلول، توانایی پاسخ به استروژن را از دست میدهند. این سلولها همچنان میتوانند با دور زدن نیاز به سیگنال رشد خارجی تولیدمثل کنند.
- سلولهای سرطانی مهار تماسی را نشان نمیدهند. در حالی که بیشتر سلولها میتوانند تشخیص دهند که توسط سلولهای مجاور محاصره شدهاند، سلولهای سرطانی دیگر به این سیگنال توقف پاسخ نمیدهند. ادامه رشد منجر به انباشته شدن سلولها و تشکیل توده تومور میشود.
- سلولهای سرطانی میتوانند بدون دریافت سیگنال «تأیید صحت همانندسازی» تقسیم شوند. در حالی که سلولهای طبیعی در حضور آسیب ژنتیکی (DNA) تقسیم را متوقف میکنند، سلولهای سرطانی به تقسیم خود ادامه میدهند. نتایج حاصل از این امر سلولهای دختری است که حاوی DNA غیرطبیعی یا حتی تعداد غیرطبیعی کروموزومها هستند. این سلولهای جهشیافته حتی غیرطبیعیتر از سلولهای «والد» هستند. به این ترتیب، سلولهای سرطانی میتوانند تکاملیافته و به تدریج غیر طبیعیتر شوند.
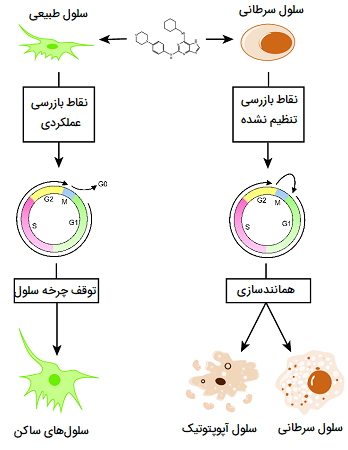
در این تصویر تفاوت بین سلول نرمال و سرطانی در ادامه همانندسازی نشان داده شده است.

ادامه تقسیم سلولی منجر به تشکیل تومورها میشود. بیثباتی ژنتیکی ناشی از تقسیم نابجا به مقاومت دارویی که در بسیاری از سرطانها دیده میشود کمک میکند. جهش در ژنهای خاص میتواند رفتار سلولها را به گونهای تغییر دهد که منجر به افزایش رشد یا توسعه تومور شود.
آزمون سنجش یادگیری
۱. مهمترین تفاوت ساختاری تقسیم میتوز (Mitosis) و شکافت دوتایی (Binary Fission) با توجه به اجزای سلولی درگیر چیست؟
در شکافت دوتایی تکثیر DNA و تقسیم کاملا همزمان است اما در میتوز جداگانه انجام میشود.
در هر دو فرآیند، غشای هستهای حفظ میشود و تغییر نمیکند.
در میتوز جداسازی کروموزومها توسط پلاسمیدها صورت میگیرد ولی در شکافت دوتایی توسط میکروتوبولها
در میتوز ساختار دوک میتوزی تشکیل میشود اما شکافت دوتایی فاقد این ساختار است.
در تقسیم میتوز، ساختار دوک میتوزی نقش مهمی در جداسازی کروموزومها دارد، ولی در شکافت دوتایی باکتریایی این ساختار وجود ندارد و تقسیم سادهتر انجام میشود. برخلاف عبارت «غشای هستهای حفظ میشود»، در میتوز غشای هستهای از بین میرود. عبارت «تکثیر DNA و تقسیم همزمان» درباره همانندسازی DNA صحیح است اما به عنوان تفاوت اصلی ساختاری مطرح نیست. همچنین، وظیفه جداسازی کروموزومها در میتوز برعهده دوک و میکروتوبولهاست، نه پلاسمیدها.
۲. کدام بخش چرخه سلولی بیشترین زمان را پیش از تقسیم واقعی سلول به خود اختصاص میدهد و هدف اصلی این بخش چیست؟
تلوفاز؛ بازسازی پوشش هستهای
متافاز؛ آرایش کروموزومها در وسط سلول
پروفاز؛ تبدیل کروماتین به کروموزوم
اینترفاز؛ رشد سلول و تکرار DNA
بخش «اینترفاز» بیشترین زمان چرخه سلولی را پیش از آغاز تقسیم به خود اختصاص میدهد زیرا طی آن سلول رشد میکند، مواد مغذی و انرژی ذخیره مینماید و مهمتر اینکه تکرار DNA و تکثیر اندامکها انجام میشود. مراحل پروفاز، متافاز و تلوفاز، همگی مربوط به تقسیم واقعی و کوتاهتر از اینترفاز هستند. بنابراین، هدف اساسی اینترفاز، آمادهسازی کامل سلول برای تقسیم سالم است که با رشد، همانندسازی DNA و افزایش اجزاء سلولی تحقق مییابد.
۳. اگر عملکرد پروتئین p53 در یک سلول انسانی مختل شود، چه اثری بر چرخه تقسیم سلولی دارد؟
سلول توانایی عبور از نقطه بازرسی G2 را از دست میدهد.
سلول در مرحله اینترفاز باقی میماند و تقسیم نمیشود.
تنوع ژنتیکی سلولهای تولیدی کاهش مییابد.
تقسیم سلولی حتی با وجود آسیب DNA ادامه مییابد.
پروتئین p53 در شرایط آسیب DNA تقسیم سلولی را متوقف میکند تا یا سلول ترمیم شود یا به حذف برنامهریزیشده برود. در صورت نقص این پروتئین، سلول حتی در صورت وجود خطاهای ژنتیکی به تقسیم ادامه میدهد. بنابراین، تنها «تقسیم سلولی حتی با وجود آسیب DNA ادامه مییابد» درست است؛ زیرا گزینههای دیگر اثری مربوط به نقش مستقیم p53 در ایست تقسیم یا کاهش تنوع ژنتیکی ندارند. باقی ماندن در اینترفاز به عملکرد صحیح p53 بستگی ندارد و از دست دادن عبور از نقطه بازرسی G2 بیشتر وابسته به کنترلهای دیگر است.
۴. برای تولید گامتهای هاپلوئید با تنوع ژنتیکی بالا در ارگانیسمهای یوکاریوت، کدام ویژگی فرایند میوز اهمیت اساسی دارد؟
کروموزومها به طور تصادفی در متافاز I آرایش مییابند و کراسینگاور رخ میدهد.
دوک میتوزی به طور کامل تشکیل و باعث تراز دقیق کروموزومها میشود.
پس از پایان سیتوکینز دو سلول دیپلوئید مستقل تشکیل میشود.
در پروفاز I کروموزومها فشرده میشوند و هسته سلول از بین میرود.
تنوع ژنتیکی بالا در گامتها نتیجه فرایندهایی مانند «آرایش تصادفی کروموزومها در متافاز I» و «کراسینگاور» در میوز است که باعث ترکیبهای ژنتیکی متنوع میگردد. تشکیل دوک میتوزی و آرایش دقیق کروموزومها در مرحله متافاز اهمیت حیاتی به صورت انحصاری برای تراز و جداسازی صحیح دارند اما به تنهایی عامل تنوع بالا نیستند. فشرده شدن کروموزومها و ناپدید شدن هسته در پروفاز I مقدمه تقسیم است و خودش تنوع ژنتیکی ایجاد نمیکند. تشکیل دو سلول دیپلوئید در پایان سیتوکینز به میوز مربوط نیست، چون میوز به تولید چهار سلول هاپلوئید میانجامد نه دو سلول دیپلوئید. بنابراین نقش کلیدی در تنوع ژنتیکی، همان آرایش تصادفی و کراسینگاور است که در میوز رخ میدهد.
۵. از نظر فرایند ترمیم بافت، چه تفاوتی میان تقسیم سلولی در سلولهای بنیادی و سلولهای بالغ وجود دارد؟
تقسیم سلولهای بالغ منجر به تنوع سلولی زیاد میشود و سلولهای بنیادی فقط سلول مشابه خود تولید میکنند.
هر دو نوع سلول تنها در شرایط آسیب تقسیم میشوند و الگوی تکثیر مشابهی دارند.
سلولهای بنیادی قابلیت تقسیم مکرر و جایگزینی انواع سلول را دارند ولی سلولهای بالغ اغلب تقسیم محدودی انجام میدهند.
سلولهای بالغ معمولا تقسیم دیگر سلولها را هدایت میکنند درحالیکه سلولهای بنیادی این نقش را ندارند.
در ترمیم بافت، «سلولهای بنیادی» با توانایی تقسیم مداوم و تولید انواع سلولهای تخصصی نقش اصلی را در جایگزینی سلولهای جدید ایفا میکنند، در حالی که «سلولهای بالغ» معمولا توانایی تقسیم محدود دارند و فقط در برخی بافتها میتوانند جایگزین سلولهای همنوع خود شوند. بنابراین، قدرت تجدید و ترمیم گسترده عمدتا به سلولهای بنیادی مربوط است و تقسیم سلولهای بالغ معمولا محدود به حفظ تعداد سلولهای همان نوع باقی میماند. برخلاف گفتههایی چون «سلولهای بالغ تقسیم دیگر سلولها را هدایت میکنند» یا «تقسیم سلولهای بالغ منجر به تنوع سلولی زیاد میشود»، تقسیم سلولهای بالغ معمولا باعث ایجاد سلولهایی مشابه خود میشود و نقش تنوع عمدتا بر عهده سلولهای بنیادی است؛ همچنین هر دو نوع سلول ممکن است مستقل از شرایط آسیب نیز تقسیم داشته باشند.
۶. اگر سیتوکینز در سلول گیاهی کامل انجام نشود، کدام تفاوت عمده با تقسیم سلول جانوری آشکار میشود؟
سلول گیاهی دارای شیار تقسیم همانند جانوران میشود.
سلول گیاهی فاقد دیواره سلولی جدید خواهد بود.
اندامکها به طور مساوی در دو سلول تقسیم نمیشوند.
هستههای سلول گیاهی به هم متصل باقی میمانند.
در سلول گیاهی، در صورت اختلال در سیتوکینز، ساختار فراگموپلاست و صفحه تقسیم به درستی تشکیل نمیشود، لذا دیواره سلولی جدید که دو سلول را جدا کند ایجاد نخواهد شد. این تفاوت بزرگ با سلول جانوری است که در آن شیار تقسیم توسط حلقه اکتین-میوزین سلول را جدا میکند. گزینه «سلول گیاهی فاقد دیواره سلولی جدید خواهد بود» صحیح است، زیرا سلول جانوری اصولا دیواره سلولی ندارد و تقسیم با شیار اتفاق میافتد. سایر گزینهها صحیح نیستند؛ شیار تقسیم مخصوص سلول جانوری است و اندامکها معمولا طبق سازماندهی طبیعی تقسیم میشوند. اتصال هستهها نیز جزو ویژگی اختلال در گیاهان با دیواره تشکیلنشده نیست و بیشتر به سلولهای چندهستهای مرتبط است.
۷. در شکافت دوتایی باکتری، پلاسمیدها و ریبوزومها چه نقشی دارند و این نقش چه اثری بر سلولهای دختری میگذارد؟
فقط ریبوزومها بین سلولهای دختری تقسیم میشوند و پلاسمیدها جابجا نمیشوند.
پلاسمیدها ژنهای مقاومت به آنتیبیوتیک را کپی میکنند و ریبوزومها برای تامین نیازهای متابولیکی سلولهای جدید تکثیر میشوند.
پلاسمیدها باعث تقسیم نابرابر سیتوپلاسم و ریبوزومها فقط در سلول والد باقی میمانند.
پلاسمیدها مواد مغذی را ذخیره میکنند و ریبوزومها شکنندهتر از کروموزوم هستند.
در فرآیند شکافت دوتایی، پلاسمیدها که حلقههای کوچک DNA هستند همانند کروموزوم باکتری کپی و به هر سلول دختری منتقل میشوند و از این طریق ژنهایی مانند مقاومت به آنتیبیوتیک بین سلولهای بعدی نیز باقی میماند. ریبوزومها نیز پیش از تقسیم تکثیر شده و این امر تامین نیازهای متابولیکی و ساخت پروتئین هر سلول دختری را تضمین میکند؛ «پلاسمیدها ژنهای مقاومت به آنتیبیوتیک را کپی میکنند و ریبوزومها برای تامین نیازهای متابولیکی سلولهای جدید تکثیر میشوند» دقیقا مطابق متن است. سه گزینه دیگر با محتوای متن همخوانی ندارند؛ در واقع، پلاسمیدها مواد مغذی ذخیره نمیکنند، تقسیم نابرابر انجام نمیدهند و هم پلاسمید هم ریبوزوم بین هر دو سلول دختری توزیع میگردد.
۸. چرا وجود نقاط بازرسی چرخه سلولی در جلوگیری از تومورهای سرطانی اهمیت دارد؟
امکان حذف سلولهای آسیبدیده را فراهم میکند.
باعث افزایش تقسیم سلولی سرطانی میشود.
هسته سلولها را دو برابر میکند.
موجب تسریع کراسینگاور کروموزومها میشود.
نقاط بازرسی چرخه سلولی این امکان را به سلول میدهد که اشتباهات یا آسیبهای ژنتیکی را قبل از ادامه تقسیم شناسایی کند و سلولهای مشکلدار را حذف یا متوقف کند. عدم وجود این نقاط بازرسی، باعث تقسیم بدون کنترل سلول میشود که زمینهساز رشد تومورهای سرطانی است. دیگر گزینهها نادرستاند، زیرا نه افزایش تقسیم سرطانی، نه دو برابر شدن هسته و نه تسریع کراسینگاور دلیل اهمیت نقاط بازرسی نیستند.
۹. در مرحله متافاز میتوز، چرا همتراز شدن کروموزومها روی صفحه متافازی اهمیت دارد؟
این وضعیت باعث جداسازی برابر کروموزومها به دو قطب سلول میشود.
سبب متراکم شدن بیشتر کروموزومها برای سلول جدید میگردد.
کروموزومها را از آسیب دیدگی توسط اندامکها محافظت میکند.
باعث مهار تعامل پروتئینهای سانتریول با DNA میگردد.
در متافاز میتوز، همتراز بودن کروموزومها روی صفحه متافازی اهمیت دارد چون این آرایش باعث میشود هر مجموعه کروماتید خواهری به طور برابر و صحیح به دو قطب سلول حرکت کند. به این ترتیب، اطلاعات ژنتیکی بدون نقص یا کاهش به سلولهای دختری منتقل میشود. انتخاب «جداسازی برابر کروموزومها به دو قطب سلول» صحیح است چون قرارگیری صحیح روی صفحه متافازی تضمین میکند کروموزومها بهدرستی تقسیم شوند. سایر گزینهها مانند «محافظت از کروموزومها توسط اندامکها»، «متراکم شدن بیشتر کروموزومها» یا «مهار تعامل پروتئینهای سانتریول با DNA» ربطی به هدف اصلی آرایش کروموزومها در متافاز ندارند.
۱۰. در تقسیم سلولی چرا میوز دو مرحله متوالی دارد و پیامد ژنتیکی آن با میتوز متفاوت است؟
چون میوز برخلاف میتوز موجب بازسازی بافتها میشود.
چون در میوز همانندسازی DNA قبل از هر مرحله انجام میشود.
چون میوز باید تعداد کروموزومها را کاهش دهد و تنوع ژنتیکی ایجاد کند.
چون فقط میوز اطلاعات ژنتیکی را به کلی حذف میکند.
در میوز، به دلیل تولید گامتهای هاپلوئید و نیاز به نصفشدن تعداد کروموزومها، دو مرحله متوالی از تقسیم هستهای وجود دارد. همچنین فرایندهایی مانند کراسینگاور و جداسازی تصادفی کروموزومها باعث تنوع ژنتیکی در سلولهای حاصل میشود. این تفاوت مهم است، زیرا میتوز سلولهای مشابه مادر تولید میکند و فقط یک مرحله تقسیم دارد، درحالیکه میوز برای تولید سلولهای جنسی با ترکیب ژنتیکی منحصربهفرد و نصف کروموزومها ضروری است. بنابراین، «میوز باید تعداد کروموزومها را کاهش دهد و تنوع ژنتیکی ایجاد کند» صحیح است و سایر گزینهها نادرستاند چون اشارهای به اهداف کلیدی میوز ندارند یا درباره نقش آن اشتباه بیان شدهاند.









