



تغذیه گیاهی چیست؟ – به زبان ساده + اصول و اهمیت
گیاهان گروه بزرگی از موجودات زنده خشکی و دریا هستند که بهوسیله نور خورشید، دیاکسید کربن و عناصر معدنی ترکیبات مورد نیاز برای رشد و بقای خود را میسازند. ریشه، اندامی است که وظیفه جذب آب و مواد معدنی را بر عهده دارد و برگ کارخانه تولید کربوهیدراتها است. در این مطلب انواع عناصر معدنی مورد نیاز در تغذیه گیاهی را به همراه روشهای جذب و انتقال آن از ریشه توضیح میدهیم. به علاوه با انواع کودهایی آشنا میشویم که مکملهای تغذیه گیاهی هستند.
- عناصر ضروری و نقش حیاتی آنها برای رشد گیاه را یاد میگیرید.
- خواهید آموخت چگونه مواد مغذی از طریق ریشه و برگ جذب و منتقل میشوند.
- نقش فتوسنتز در تولید مواد آلی در گیاه را یاد میگیرید.
- میآموزید عوامل محیطی مانند «pH»، دما و نوع خاک چگونه بر تغذیه گیاه اثر میگذارند.
- خواهید توانست تفاوت انواع کودها و نقش آنها در بهبود خاک را توضیح دهید.
- نقش میکروارگانیسمهای خاک در تغذیه و تثبیت نیتروژن را یاد خواهید گرفت.




تغذیه گیاهی چیست ؟
رشد و بقای گیاه به دریافت دو دسته مواد آلی و معدنی بستگی دارد. گیاه مواد معدنی مورد نیاز را از خاک دریافت میکند و بخشی از مواد آلی مورد نیاز (کربوهیدرات) را با استفاده از گاز دیاکسیدکربن و در فرایند فتوسنتز تولید میکند.
سنتز مواد آلی
فتوسنتز فرایندی است که در تمام گیاهان سبز، بعضی قارچها و باکتریها برای سنتز مواد آلی استفاده میشود. این فرایند چهارمرحلهای در بخشهای مختلف یاخته گیاهی انجام میشود و با استفاده از انرژی نور خورشید، انرژی حاصل از انتقال الکترونها و تثبیت کربن، کربوهیدراتهای مورد نیاز گیاه را میسازد.

- جذب نور: اولین مرحله این فرایند جذب نور بهوسیله کلروفیلها موجود در غشای تیلاکوئیدی (کیسههای غشایی کلروپلاست) است. جذب نور منجر به تغییر انرژی الکترون موجود در مرکز واکنش فتوسنتزی و خروج الکترون از این پروتئین میشود.
- انتقال الکترون: توالی از پروتئینهای عرض غشایی در غشای تیلاکوئیدی قرار دارند و الکترون آزاد شده از فتوسیستم II پس از انتقال بین آنها به «مولکول پذیرنده الکترون» منتقل و مولکول در واکنش با یون هیدروژن به NADPH تبدیل میشود. همزمان انرژی حاصل از انتقال الکترون، صرف انتقال پروتونها به لومن تیلاکوئید میشود.
- سنتز ATP: پمپ غشای تیلاکوئید با انتقال پروتونها از لومن تیلاکوئید به استرومای کلروپلاست، اتصال فسفات به ADP و تشکیل ATP را کاتالیز میکند. این مرحله از واکنشهای تیلاکوئیدی مشابه تولید ATP در زنجیره انتقال الکترون در میتوکندری جانوران است.
- تثبیت کربن: NADPH و ATP تولید شده در واکنشهای زنجیره انتقال الکترون، الکترون و انرژی لازم برای کاهش به کربوهیدرات پنجکربنه ریبولوز بی فسفات را تامین میکنند. نور عامل کلیدی واکنشهای سه مرحله قبل است (واکنشهای وابسته به نور). اما این مرحله از فتوسنتز بدون حضور نور و در تاریکی انجام میشود (واکنش مستقل از نور).
جذب دی اکسید کربن در برگ
دیاکسید کربن مولکول اصلی در تغذیه گیاهی از مواد آلی است. در بخش قبل توضیح دادیم که با تثبیت این گاز در چرخه کالوین کربوهیدرات لازم برای رشد گیاهان و مولکولهای حدواسط سنتز سایر ترکیبات آلی (آمینواسیدها) را فراهم میکند. این گاز از راه روزنه یا استوماتا جذب میشود. استوما منافذ کوچک روی برگ است که بهوسیله سلولهای نگهبان ایجاد میشود. «سلولهای نگهبان» (Guard cells) یاختههای گیاهی تمایزیافته از اپیدرم به شکل هلالی (شبیه نان کروسان) هستند که فراوانی آنها در سطح زیری برگ گیاهان خشکی و سطح بالایی گیاهان شناور روی آب بیشتر است. تغییر حجم سلولهای نگهبان سبب باز و بسته شدن استوما و ورود دیاکسید کربن یا خروج آب از برگ میشود. دمای هوا، غلظت آب و دیاکسید کربن سلولهای نگهبان سه عامل محیطی کنترلکننده باز و بسته شدن روزنهها هستند.

هورمون گیاهی آبسیزیک اسید، یکی دیگر از عواملی است که جذب دیاکسید کربن از استروما را تنظیم میکند. کاهش آب موجود در سلولهای برگ سبب تولید و ترشح این هورمون میشود. اتصال آبسیزیک اسید به رسپتور آن در غشای سلولهای نگهبان با کاهش حجم سلولها، بسته شدن استوما و جلوگیری از تبخیر آب و ورود دیاکسید کربن همراه است.
تغذیه گیاه از مواد معدنی
علاوه بر کربوهیدراتهایی که در فرایند فتوسنتز تولید میشوند، گیاه نیاز به مواد معدنی خاصی دارد که فرایندهای مختلف زیستی و ترکیبات آلی دیگر را تولید کند. فسفر، نیتروژن و کلسیم، عناصر معدنی مهم مورد نیاز گیاه هستند. برخی از مواد معدنی برای رشد گیاه حیاتی هستند. گیاه بدون این ۱۵ عنصر اصلی نمیتواند رشد کند و از بین میرود. این عناصر ضروری در دو دسته «درشت مغذی» (Macronutrients) و «ریزمغذی» (Micronutrients) قرار میگیرند و نیاز گیاه به مواد درشت بیشتر از ریزمغذیها است. عناصر غیرحیاتی، گروهی از ترکیبات معدنی موردنیازی هستند که کمبود آنها منجر به بیماری یا توقف رشد گیاه نخواهد شد. این عناصر به شکل یونهای محلول در آب یا متصل به ذرات خاک وجود دارند و در زمان نیاز با مکانیسمهای مختلف تبادل یونی از ریشه وارد گیاه میشوند.
درشت مغذی ها
درشت مغذیها شامل عناصر کربن، هیدروژن، نیتروژن، فسفر، پتاسیم، کلسیم، گوگرد، منیزیوم (برای سنتز کلروفیل) و آهن میشود که هر کدام نقش مهمی در متابولیسم و سنتز گیاه دارند و کمبود آنها با ایجاد بیماری در گیاه همراه است.
- نیتروژن: عنصر حیاتی برای سنتز آمینواسیدها، DNA، هورمونهای گیاهی و کلروفیل در گیاهان است. یون نیترات شکل قابل جذب نیتروژن جو، نیتروژن معدنی و نیتروژن آلی برای گیاه است. کمبود این عنصر در گیاه منجر به دوکی، زرد شدن و در بعضی موارد تشکیل دانههای صورتی روی برگ میشود.
- فسفر: برای انجام تقسیم سلولی و رشد بخشهای مختلف گیاه ضروری است. این عنصر در ساختار بسیاری از مولکولهای زیستی (نوکلئوتیدها، آمینواسیدها، پروتئینهای تنظیمی و آنزیمها) فرایند تقسیم وجود دارد. کمبود فسفر منجر به رشد کند گیاه و زرد و شکننده شدن برگها میشود.
- کلسیم: عنصر ضروری برای رشد ریشه، تشکیل تارهای کشنده و رشد برگ گیاهان است.
- منیزیوم: عنصر کلیدی ساختار کلروفیل و تداوم واکنشهای فتوسنتزی است. بدون حضور این عنصر کلروفیلی تشکیل نمیشود. در نتیجه فتوسنتز و رشد گیاه متوقف خواهد شد. کمبود این عنصر منجر به زرد شدن برگ در اطراف رگبرگها میشود که در بعضی گیاهان با تشکیل لکههای قرمز مایل به قهوهای و ریزش زود هنگام برگها همراه است.
- پتاسیم: به انتقال مولکولهای قند، چربیها و پلیمر نشاسته در گیاه کمک میکند.
- گوگرد یکی دیگر از عناصر ضروری رشد گیاه است که در سنتز آمینواسیدها شرکت میکند. به علاوه تشکیل بسیاری از مولکولهای بو و مزه در گیاهان (برای مثال در پیاز و کلم) به وجود این عنصر بستگی دارد. کمبود این عنصر در خاک منجر به ایجاد لکههای زرد یا بنفش روی برگ و قهوهای شدن لبههای برگ میشود. در این شرایط تشکیل گل و میوه در گیاه کاهش مییابد.
- آهن: یکی از عناصر مهم ساختار سیتوکرومهای زنجیره انتقال الکترون و بسیاری از متالوآنزیمها است. این زنجیرهها با تامین ATP لازم برای انجام واکنشهای زیستی و انجام فتوسنتز نقش حیاتی در رشد و متابولیسم گیاهان دارد.
ریزمغذی ها و عناصر غیر حیاتی
منگنز، روی، بورون، مس، مولبیدیوم و کبالت عناصر مهمی در تغذیه گیاهی هستند که نیاز گیاه به انها کمتر از عناصر درشتمغذی است.
- منگنز: موجود در فتوسیستم II تیلاکوئیدی به هیدرولیز نوری آب کمک میکند. کمبود منگنز و آهن علائم مشابهی در برگ گیاه ایجاد میکند. کمبود این دو عنصر فلزی منجر به زرد شدن برگها در اطراف رگبرگها و قهوهای شدن لبههای برگ در گیاهان اسیددوست میشود.
- مس: یکی دیگر از عناصر موجود در متالوآنزیمهای گیاهی است.
- روی عنصری است که در مسیرهای سنتز هورمونهای مربوط به افزایش رشد طولی و عرضی ساقه و برگها شرکت میکند. غلظت این عنصر در خاکهای اسید بیشتر است اما تمایل زیادی به برهم کنش با یون آهن دارد.
- بورون: یکی از عناصر لازم برای حفظ و پایداری شکل سلولها در گیاهان آوندی است. این عنصر در تشکیل اتصالات عرضی پکتینهای رامنوگالاکتورونان II در دیواره سلولی شرکت میکند.
- مولبیدیوم: عنصر ضروری برای سنتز آنزیم نیتروژناز در میکرواورگانیسمهای تثبیتکننده نیتروژن است. به همین دلیل خاک مناسب برای کشت حبوبات باید مولبیدیوم کافی داشته باشد. به علاوه سلول گیاهی برای سنتز آمینواسیدها از نیتروژن محلول به این عنصر (متالوآنزیم) نیاز دارد. کمبود این عنصر در گیاهان خانواده کلم منجر به تشکیل برگهای طویل و پیچخورده میشود.
- عناصر غیرحیاتی: سدیم، آلومینیوم، سیلیکون، کلر و گالیوم عناصر غیرحیاتی هستند که نیاز گیاه به آنها بسیار کمتر است.

ساختارهای جذب مواد معدنی در گیاه
گیاهان آبی برای تغذیه از مواد معدنی نیاز به ساختار ویژهای ندارند. در این گیاهان تمام بدن جاندار سطحی برای دریافت مواد معدنی و تبادل یونها با محیط آبی اطراف است. بیشتر این تبادل به دلیل اختلاف فشار اسمزی و غلظت یونها در آب و یاختههای گیاهی انجام میشود. در گیاهان خشکی ریشه ساختار اصلی است که با خاک در ارتباط است و تبادل یونها و عناصر معدنی را بر عهده دارد. مریستمها، سلولهای فعال گیاهی هستند که ویژگی تقسیم سلولی در آنها حفظ شده است. فعالیت متابولیکی این بخش ریشه بیشتر از سایر بخشها است و بیشتر جذب مواد معدنی را بر عهده دارد. عناصر جذب شده با انتقال فعال از سلولهای ریشه به آوند چوبی و بهوسیله این ساختارها به سایر بخشهای گیاه منتقل میشوند.
ساختار ریشه
به طور کلی ریشه بخشی از ساختارهای گیاهی است که در خاک قرار دارد و مورفولوژی آن بر اساس نوع گیاه (دولپه و تکلپهای، گلدار و بدون گل) و منطقه زندگی آن متفاوت است. این بخش از گیاه علاوه بر جذب مواد معدنی مثل لنگری به باقی ماندن گیاه در خاک و استاده ماندن ساقه کمک میکند. ریشه از سه بخش مریستمی، بخش تمایزی و بخش افزایش طول تشکیل شده است.
- مریستم ریشه: از سلولهای تمایزنیافته مریستم راسی تشکیل میشود که به سرعت بهوسیله تقسیم میتوز تکثیر میشوند و سلولهای جدید برای رشد ریشه را ایجاد میکنند.
- بخش افزایش طول: طول سلولهای حاصل از تقسیم میتوز پس از ورود به این افزایش یافته و به افزایش طول ریشه و نفوذ بیشتر در خاک کمک میکنند.
- بخش تمایزی: این بخش از سلولهای بالغ و تمایزیافتهای تشکگیل شده است که آوند چوبی، آبکش و تار کشنده را میسازند.
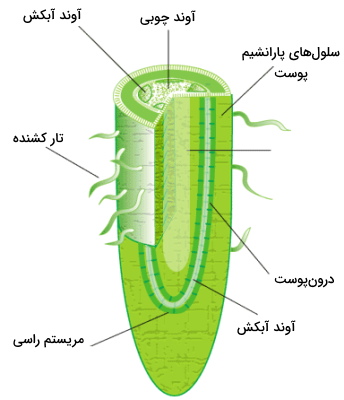
ریشه مثل سایر بخشهای گیاه از بافتهای مختلفی تشکیل شده است. اگر برش عرضی این ساختار را در نظر بگیریم، خارجیترین بخش لایهای از ماده مومی کوتیکول است که از گیاه در برابر تبخیر آب و آسیب آفتها (عفونتهای باکتریایی و قارچی) محافظت میکند. البته برای جذب آب، این لایه در ریشه نسبت به برگگیاهان بسیار نازکتر است. پس از این لایه مومی (نوعی از مولکولهای چربی) روپوست، پوس، درونپوست، نوار کاسپاری و استوانههای آوندی (مجموعهای از پریسایکل، آوند چوبی و آوند آبکش با آرایش متفاوت در گیاهخان دولپهای و تکلپهای) قرار دارند.
- روپوست: اپیدرم یا روپوست خارجیترین لایه بافتی ریشه است که از لایههای داخلی در برابر آسیبهای محیطی محافظت میکند. روپوست ریشه از یک لایه سلول تشکل میشود.
- پوست: پوست یا کرتکس بخشی چند لایه در ریشه و وظیفه آن ذخیره آب و مواد معدنی است.
- درونپوست: اندودروم یا درونپوست در ریشه از یک ردیف سلول گیاهی تشکیل میشود. ضخامت دیواره سلولهای این لایه به دلیل وجود پلیساکاریدهای لیگنین و سوبرین از سلولهای پوست بیشتر است. این پلیساکاریدها سدی ضد آب به نام نوار کاسپاری در درونپوست میسازند که از جریان سیتوپلاسمی آب جلوگیری میکند.
- پریسایکل: پریسایکلها اولین سلولهای دستهجات آوندی از مرکز برش عرضی ریشه هستند که وظیف اصلی آنها تولید ریشههای عرضی است.
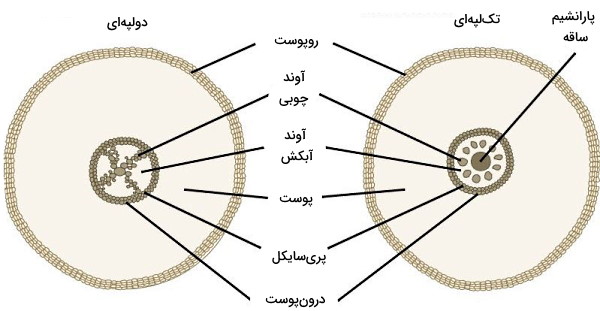
- آوند چوبی: آوندهای چوبی یا زایلمها ساختارهای لولههای انتقال آب و مواد معدنی در گیاهان آوندی هستند. آرایش این آوندها در ریشه گیاه جوان و دولپهای به شکل حرف x انگلیسی و در گیاهان تکلپهای به شکل حلقههایی اطراف مرکز است.
- آوند آبکش: این آوندها لولههای انتقال کربوهیدراتهای تغذیه گیاهی هستند و در ریشه معمولا بین آوندهای چوبی قرار میگیرند.
تار کشنده
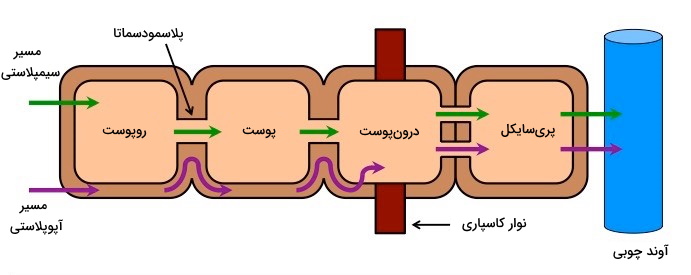
تار کشنده زوائد اپیدرمی ریشه هستند و یکی از مراکز تبادل آب و مواد معدنی با خاک است. این زوائد قطر بسیار کم و مساحت سطح بسیار زیادی دارند که بهترین ویژگی برای تبادل مواد بر اساس اختلاف فشار اسمزی سیتوپلاسم سلولها و خاک را فراهم میکند. آکوآپورینهای موجود در غشای سلولهای تارکشنده به ورود آب کمک میکنند. آب در عرض ریشه تا آوند چوبی بر اساس اختلاف شیب غلظت و از مسیر آپوپلاستی یا سیمپلاستی حرکت میکند.
- مسیر آپوپلاستی: در این مسیر آب از بین دیواره و فضای بین سلولی (فضای آزاد در دسترس) عبور میکند. در این مسیر آب وارد سیتوپلاسم سلولهای کناری نخواهد شد. آب و مواد معدنی محلول در آن در این روش بدون نیاز به ATP و غیرفعال منتقل میشوند. ترکیبات باردار موجود در دیواره سلولی به حرکت سریع یونها از راس تار کشنده تا درونپوست کمک می کنند. اما در درونپوست نوار کاسپارینی مثل سدی محکم اجازه عبور آب را نمیدهد و حرکت آب تنها از پلاسمودسماتای بین سلولی و با ورود به سیتوپلاسم ادامه مییابد.
- مسیر سیمپلاستی: در مسیر سیمپلاستی مولکولهای آب از پلاسمودسما وارد سیتوپلاسم سلول شده (اما وارد واکوئل نمیشود) و به سلول کناری منتقل میشود. از آنجایی که آب برای رسیدن به آوندهای چوبی در این مسیر از غشای پلاسمایی عبور میکند، به آن جریان سیتوپلاسمی نیز گفته میشود.
مکانیسم های گیاهی جذب مواد معدنی
یونها و عناصر معدنی از دو مسیر فعال و غیرفعال وارد سلولهای ریشه و بین این سلولها منتقل میشوند. تبادل یونی، تعادل دونان و جریان تودهای مکانیسمهایی هستند که به انتقال غیرفعال یونها از ریشه کمک میکنند. انتقال فعال این یونها مثل غشای سلولهای جانوری به ناقلهای پروتئینی عرض غشایی نیاز دارد.
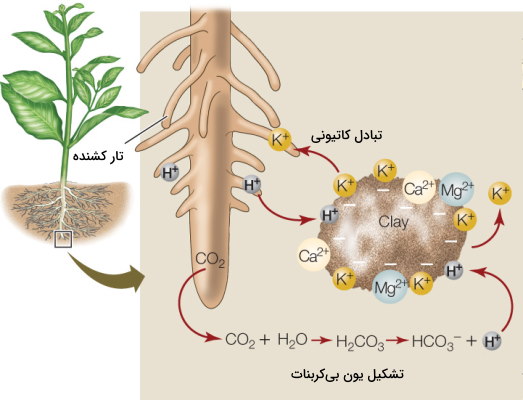
تبادل یونی مکانیسمی است که انرژی مورد نیاز برای ورود یونهای معدنی به ریشه را بهوسیله انرژی حاصل از اختلاف پتانسیل یونی تامین میکند. این مکانیسم از یونهای هیدروژن و بیکربنات برای انتقال کاتیونهای مختلف به ریشه بهره میبرد. بخش زیادی از یونها با ذرات رس موجود در خاک ترکیبات کلوئیدی تشکیل میدهند و مکانیسم ورود آنها به سلولهای گیاهی ریشه با مولکولهای آب متفاوت است. در غشای سلولهای روپوست ریشه پمپهای هیدروژنی وجود دارد که با مصرف انرژی یون هیدروژن ذخیره شده در واکوئلها را از سلول خارج و به خاک منتقل میکنند. یون هیدروژن جایگزین کاتیونها (، ، ، و ) در کلوئید خاک شده و کاتیونها در جهت شیب غلظت وارد سیتوپلاسم سلولهای ریشه میشوند. ظرفیت نگهداری برای کاتیونهای هیدروژن در ذرات رس (با بار منفی) از همه بیشتر و ظرفیت نگهداری کاتیون سدیم از همه کمتر است. به همین دلیل هیدروژن میتواند جایگزین تمام کاتیونها شود.
به علاوه دیاکسید کربن حاصل از تنفس سلولی ریشه پس از ورود به خاک با مولکولهای آب اطراف واکنش میدهد و کربونیکاسید تشکیل میشود. از تجزیه سریع این اسید ضعیف و ناپدار یون هیدروژن و بیکربنات تولید خواهد شد. کاتیونهای کلوئیدی ممکن است با یون هیدروژن مبادله یا پس از اتصال به بیکربنات وارد ریشه شوند.
نیتروژن و گوگرد دو عنصر حیاتی دیگر هستند که در سنتز آمینواسیدها نقش دارند. نیترات ()، آمونیوم () و سولفات () موجود در خاک مهمترین منابع دریافت این دو عنصر هستند که ترکیب آنها در بخشهای مختلف خاک و بر اساس شرایط آب و هوا تغییر میکند. نیترات جذب شده در سلولهای ریشه برای شرکت در سنتز آمینواسیدها به آمونیاک تبدیل میشود. این واکنش با تولید یون هیدروکسید (OH) همراه است که باید به خاک منتقل یا با اسیدهای آلی خنثی شود. به همین دلیل در زمان جذب نیترات، محیط اطراف ریشه قلیایی است. برای حفظ تعادل الکتروشیمیایی ورود نیترات با خروج آنیونها یا ورود کاتیون به سلول همراه شود. برای مثال گیاه گوجهفرنگی نیترات را همراه با یکی از یونهای فلزی پتاسیم، کلسیم، منیزیوم یا سدیم جذب میکند و این یون را به شکل اسیدهای آلی مالات و اگزالات ذخیره میکند. اما در گیاه سویا همزمان با ورود نیترات به سلول، هیدروکسید یا بیکربنات () از سلول وارد خاک میشوند.
کلر () و بوروم () دو یون دیگر مورد نیاز گیاه هستند که در تنظیم فشار اسمزی سلولها نقش دارند. این آنیونها بر اساس اختلاف شیب پتانسیل الکتریکی مبادله میشوند. این انتقال تا زمانی ادامه دارد که پتانسیل بار الکتریکی در دو طرف غشای پلاسمایی یکسان شود. در این حالت نوع یونهای موجود در دو طرف غشا یکسان نیست. اما جمع جبری بار منفی و مثبت دو طرف غشا با هم برابر است (تعادل دونان).
بر اساس نظریه جریان تودهای (یکی از مکانیسمهای انتقال مواد در آوند چوبی) تبخیر آب در فرایند تعریق از برگ سبب جذب بیشتر آب همراه با یونها محلول از ریشه میشود. در نتیجه افزایش تعریق از روزنههای برگ در هوای گرم، جذب یونها و عناصر معدنی گیاه را افزایش میدهد. این مکانیسم بر اساس انتشار ساده و بدون نیاز به مصرف ATP انجام میشود.
غشای پلاسمایی سلولهای تار کشنده به همه یونها نفوذپذیر نیست و یونها به کمک ناقلهای پروتئینی متفاوت که از انرژی ATP استفاده میکنند یا بهوسیله انتشار تسهیل شده وارد سیتوپلاسم سلول خواهند شد. «اثر تجمعی یون» (Salt Accumulation) یا اثر اشباع یکی از مکانیسمهایی است که در انتقال فعال مواد معدنی به سلولهای ریشه را نقش دارد. غشای سلولهای مریستمی در راس ریشه نفوذپذیری زیادی به یونها دارد. در نتیجه یونها بر اساس شیب غلظت وارد این سلولها میشوند. پس از مدتی غلظت یونهای در سیتوپلاسم و واکوئلهای آبی این سلولها از غلظت یونها در خاک بیشتر خواهد شد. به همین دلیل برای انتقال یونهای بیشتر به سیتوپلاسم و تامین مواد معدنی کافی برای بخشهای مختلف گیاه، از انرژی و انتقال فعال استفاده کند. تنفس سلولی در میتوکندری یاختههای مریستمی فرایندی است که ATP لازم برای انتقال یونها را فراهم میکند.
یکی از این ناقلهای پروتئینی سیتوکروم موجود در زنجیره انتقال الکترون است. این تئوری اولین بار توسط اچ. لاندرگارد (H. Lundegard) مطرح شد. طبق نظر این دانشمند سیتوکرومهای موجود در غشای سلول به انتقال آنیونها کمک ميکنند و انرژی لازم برای این انتقال با اکسایش حدواسطهای تنفس سلولی تامین میشود. در این شرایط انتقال آنیونها از خاک به سلول سبب افزایش تنفس سلولی میشود و به آن تنفس آنیونی یا نمکی میگویند.
عوامل موثر بر تغذیه گیاهی از خاک
عوامل مختلفی تغذیه گیاهی از خاک و دریافت مواد معدنی را کنترل میکنند. هوازدگی خاک، دما، pH خاک، غلظت یونها و عناصر در خاک و برهمکنش بین یونها ازجمله عوامل کنترلکننده این فرایند هستند.

هوازدگی خاک
ریشه مثل سایر بخشهای گیاه از سلولهای زندهای تشکیل شده است که برای ادامه حیات و تولید انرژی لازم برای انجام فرایندهای سنتزی و انتقال مواد به اکسیژن نیاز دارند. فشردگی خاک با خاک رس و پر شدن منافذ بین ذرات خاک با آب، میزان هوای در دسترس ریشه را کاهش میدهد. در نتیجه اکسیژن کافی برای تولید ATP وجود ندارد و انتقال عناصر معدنی کاهش مییابد.
دما
تغییر دمای خاک متابولیسم سلولهای ریشه و حرکت یونها را تغییر میدهد. با کاهش دما، جذب مواد معدنی به شدت کاهش و با افزایش دما (تا یک بازه مشخص) جذب این عناصر افزایش مییابد. تغییر شدید میران جذب عناصر معدنی و یونها بهوسیله تغییر دما نشان میدهد انتقال این مواد به آنزیمها و پروتئینها وابسته است.
غلظت یون هیدروژن
تبادل بسیاری از کاتیونها بین سلولهای ریشه و خاک بهوسیله جایگزینی یون هیدروژن در کمپلکسهای خاک رس انجام میشود. به علاوه تغییرات pH شکل یون موجود در خاک را تغییر میدهد. برای مثال بیشتر یون فسفات در pHهای قلیایی به یون دوظرفیتی () و سهظرفیتی () تبدیل میشود که گیاه نمیتواند آنها را جذب کند. اما در pH خنثی یونها تکظرفیتی هستند و گیاه مکانیسمهای لازم برای جذب آنها را دارد. در نتیجه pH خاک نه تنها سرعت جذب یونها از ریشه بلکه نوع یون جذب شونده را کنترل میکند. به همین دلیل تنظیم pH خاک یکی از فاکتورهای مهم در کشاورزی است و خاک خیلی اسیدی یا خیلی قلیایی برای کشت گیاه مناسب نیست.
غلظت عناصر در خاک
به طور کلی غلظت یونها و عناصر معدنی در سیتوپلاسم سلولهای ریشه بسیار بیشتر از غلظت این مواد در خاک است که نشان میدهد بخش زیادی از این عناصر خلاف جهت شیب غلظت و با انتقال فعال وارد سلول میشوند. سرعت جذب این عناصر زمانی که آب بیشتری در خاک وجود دارد افزایش مییابد.
برهم کنش بین یونی
آنیونها و کاتیونهای موجود در خاک و حتی یونهای دارای یک نوع بار الکتریکی با هم برهمکنش دارند و ایین برهم کنشها سبب افزایش جذب هر دو یون (تسهیل جذب یون) یا کاهش جذب (اثر مخالف یون) یکی از آنها میشود. برای مثال افزایش یونهای دوظرفیتی کلسیم و منیزیوم، جذب یون آهن و پتاسیم را کاهش میدهد. به علاوه غلظت کلسیم کلرید () خاک با مهار جذب مس از مسمومیت گیاه جلوگیری میکند. اما افزایش غلظت سدیم کلرید، افزایش جذب بسیاری از یونها را به دنبال دارد. اثر مخالف یون و تسهیل جذب یون به این دلیل ایجاد میشود که ناقلهای پروتئینی مختلفی یونها را منتقل میکنند. یونهایی که برای اتصال به ناقل با هم رقابت میکنند، اثر مخالف و یونهایی که با هم منتقل میشوند، افزایش جذب دارند. در نتیجه ترکیب یونهای موجود در خاک در تغذیه گیاهی و جذب مواد معدنی بسیار مهم است.
بهبود تغذیه گیاهی از خاک
کشت و برداشت مداوم گیاهان در خاک منجر به مصرف مواد معدنی و عناصر موجود در خاک میشود. به همین در کشت بسیاری از محصولات کشاورزی، باغداری و گیاهان آپارتمانی از کودهای آلی، معدنی و میکروبی برای غنیسازی خاک و بازسازی ترکیب عناصر در آن بهره برده میشود. فرایند تولید این کودها با ایجاد تغییر در چرخه زیستی طبیعت، افزایش گازهای گلخانهای، فرسایش خاک، کاهش صخرههای فسفاتی و مصرف سوختهای فسیلی در فرایند هابر-بوش (برای تولید کودهای نیتروژن) همراه است. به همین دلیل یکی از راههای جایگزین برای بهبود خاک و تغذیه گیاهی استفاده همزمان از کودهای آلی و میکرواورگانیسمهایی است که میتوانند با ریشه گیاهان همزیستی داشته باشند و ترکیبات آلی را به عناصر معدنی مورد نیاز گیاه تبدیل کنند. مواد آلی این کودها را میتوان از پسماندهای کشاورزی، صنایع مختلف و شهری تولید کرد. به علاوه طول عمر ترکیبات معدنی در خاک بیشتر از مواد معدنی است و انحلال کمتر آنها در آب احتمال تبخیر، نفوذ به لایههای زیری خاک و خارج شدن از دسترس گیاه را کاهش میدهد.
کود شیمیایی
کودهای شیمیایی اغلب حاوی مواد غذی حیاتی برای رشد گیاه هستند کودهای نیتروژنی، کودهای فسفات، کودهای پتاسیم، کودهای ترکیبی و کودهای کامل (NPK) پنج دسته از کودهای شیمیایی هستند که روشهای تولید و ترکیب عناصر تشکیلدهنده آنها با هم متفاوت است.
- کودهای نیتروژنی: سدیم نیترات، آمونیوم سولفات، آمونیوم نیترات، آمونیوم سولفات نیترات، آمونیوم کلرید، اوره و کلسیم آمونیوم نیترات، انواع مختلف کودهای شیمیایی حاوی نیتروژن هستند.
- سدیم نیترات: ۱۶٪ ترکیب کودهای سدیم نیترات را نیتروژن موجود در یون نیترات تشکیل می دهد. این یون انحلالپذیری بسیار زیادی در آب دارد و بهراحتی با آبدهی زیاد به گیاه یا بارانهای شدید وارد لایههای زیری خاک و از دسترس ریشه خارج میشود.
- آمونیوم سولفات: حدود ۲۰٪ از ترکیب بلور سفید آمونیوم سولفات از نیتروزن تشکیل شده است. این کود برای گیاهان مناطق مرطوب ازجمله برنج و کتان بسیار مناسب است. مصرف زیاد و مداوم این کود با کاهش pH خاک (افزایش اسیدیته)، جذب برخی کاتیونها را کاهش میدهد. مصرف زیاد کود آمونیوم سولفات غلیظ کاهش جوانهزنی گیاه را به همراه دارد.
- آمونیوم نیترات: کود آمونیوم نیترات از ترکیب تقریبی ۳۵٪ نیتروژن، ۵۰٪ نیتروژن نیتراتی و ۵۰٪ آمونیوم تشکیل شده است. این کود جذب آب فراوانی دارد و به همین دلیل نگهداری آن نسبت به سایر کودهای نیتروژن سختتر است. انحلالپذیری کمتر آمونیوم نسبت به نیترات در آب سبب کاهش شستوشوی این یون به لایههای زیری خاک میشود. از آنجایی که این کود اسیدیته خاک را افزایش میدهد، قبل از اضافه شدن به خاک با آهک مخلوط میشود.
- آمونیوم سولفات نیترات: این کود مخلوطی از آمونیوم سولفات و آمونیوم نیترات است و از حدود ۲۶٪ نیتروژن (یکسوم آن به شکل آمونیاک و بقیه به شکل نیترات) تشکیل میشود. کاهش pH خاک در استفاده از کودهای آمونیوم سولفات نیترات از کودهای نیتروژن قبلی کمتر است. استفاده از این کود در مرحله رشد دانه مناسب نیست.
- آمونیوم کلرید: آمونیوم کلرید ترکیبی با ۲۶٪ نیتروژن آمونیومی است و اثری مشابه کودهای آمونیوم سولفات بر شرد گیاه دارد.
- اوره: بلور سفید اوره ترکیبی آلی است که نسبت به سایر کودهای نیتروژن بیشترین درصد این عنصر (حدود ۴۵٪ نیتروژن آلی) را دارد. این ترکیب انحلالپذیری بسیار زیادی در آب دارد و معمولا برای افزایش زمان نگهداری با یک ترکیب غیرجاذب آب پوششدار میشود. از این کود میتوان برای انواع محصولات کشاورزی بهره برد. اما استفاده از این کود در مرحله جوانه زدن به رشد گیاه آسیب میزند.
- کلسیم آمونیوم نیترات: کود کلسیم آمونیوم نیترات از ترکیب آمونیوم نیترات و سنگ آهک تولید میشود و برخلاف دیگر کودهای نیتروژنی تقریبا خنثی و مناسب خاکهای اسیدی است. عنصر هیدروژن حدود ۲۸٪ ترکیب این کود را تشکیل میدهد که ۵۰٪ آن به شکل آمونیاک و ۵۰٪ آن به شکل نیترات است.
- کود فسفات: فسفات خنثی، فسفات تغییریافته، فسفات دو محصولی و فساتهای شیمیایی انواع مختلف این کود شیمیایی هستند.
- کود پتاسیم: پتاسیم کلرید (پتاس موریات) و پتاسیم سولفات (پتاس سولفات) انواع کود پتاس یا پتاسیمی هستند که برای بهینهسازی خاک استفاده می شوند. تشکیل کلوئیدهای خاک رس یکی از ویژگیهای مهم پتاسیم است که از شسته شدن راحت این عنصر جلوگیری میکند. این ویژگی در کودهای شیمیایی این عنصر نیز وجود دارد. پتاسیم موجود در این کودها به شکل پتاس () و درصد آن در ترکیب پتاس موریات بیشتر (حدود ۵۰ تا ۶۰ ترکیب اصلی) است.
- کود ترکیبی: این کودها از ترکیب شدن دو یا چند عنصر مورد نیاز رشد گیاه تولید می شوند. برای تولید این کودها میتوان دو کود تکعنصره با نسبت مناسب ترکیب و به خاک اضافه کرد.
- کود کامل: کودهای کامل یا کودهای NPK ترکیبی از سه عنصر ضروری نیتروژن، فسفر و پتاسیم هستند که در بعضی مواقع ترکیبات ضدقارچی، نمکهای جیوه و مس و سموم ضد آفات به آنها اضافه میشود.
نقش میکرواورگانیسم ها در تغذیه گیاهی
خاک تنها یک منبع مواد معدنی و یونها برای رشد گیاه نیست. بلکه اکوسیستم پیچیدهای است که میلیونها قارچ، باکتری، ویروس و جانور آغازی در آن زندگی میکنند که متابولیسم آنها غلظت مواد معدنی موجود در دسترس گیاه را تغییر میدهد. این میکرواورگانیسمها ممکن است برای جذب مواد معدنی با گیاه رقابت کنند، بخشی از مواد معدنی و آلی مورد نیاز گیاه را تامین کنند، در گیاه ایجاد بیماری کنند یا اثری برهم نداشته باشند. این میکرواورگانیسمها بهوسیله سه مکانیسم اصلی رشد گیاه را افزایش میدهند.
- تغییر هورمونهای گیاهی
- مهار یا رقابت با سویههای پاتوژن
- افزایش مواد معدنی دسترس گیاه
بیشتر نیتروژن، فسفر و گورگد موجود در خاک به شکل ترکیبات آلی است که گیاه نمیتواند از آن استفاده کند. قارچها و باکتریهای موجود در خاک در فرایندهای متابولیسمی خود این مواد را تجزیه و عناصر N، P و S را آزاد میکنند. عناصر آزاد شده در خاک به یونهای آمونیوم، نیترات، فسفات و سولفات تبدیل و گیاه آن را جذب میکند.
علاوه بر متابولیسم ترکیبات آلی، تثبیت نیتروژن یکی از مهمترین وظایف میکرواورگانیسمهای موجود در خاک است. در این فرایند نیتروژن مولکولی () موجود در هوا به یونهای قابل جذب گیاه تبدیل میشود. باکتریهای اتوتروف ازجمله نیتروباکتر سویههایی از باکتری هستند که مواد لازم برای رشد و تقسیم سلولی خود را در فرایندهای اکسایش و بدون نیاز به موجود زنده دیگری تامین میکنند. این باکتریها برخلاف گیاهان که از انرژی فتوسنتز برای تثبیت کربن استفاده میکنند با اکسایش یون نیتریت ( NO_3^−$$) انرژی لازم برای چرخه کربس را فراهم میکنند. این فرایند در واکنش زیر انجام میشود.
سیانوباکتریها گونه دیگری از میکرواورگانیسمهای تثبیتکننده نیتروژن هستند. این میکرواورگانیسمها در شرایط معمول از نور خورشید و اکسیژن برای تامین انرژی و سنتز مواد آلی مورد نیاز خود استفاده میکنند. اکسیژن و نیتروژن در دو نوع سلول و کاملا مجزا از هم اکسید میشوند. این باکتریها در شرایط کمبود اکسیژن، نیتروزن مولکولی موجود در جو را با واکنشهای آنزیمی و با کمک نیتروژناز به نیترات تبدیل میکنند. شدت نور، غلظت نیتروژن آلی یا معدنی و دمای خاک عملکرد سیانوباکتری را تنظیم میکند.
ریزوبیومها دسته سوم میکرواورگانیسمهای مهم خاک هستند که در تثبیت نیتروژن نقش دارند. این باکتریهای گرم منفی برخلاف دو گونه دیگر هتروتروف هستند و به صورت همزیست در گرهکهای ریشه حبوبات و سایر گیاهان گلدار زندگی میکنند. ریزوبیومها با استفاده از آنزیم نیتروژناژ، نیتروژن مولکولی جو را به آمونیاک و سپس ترکیبات آلی گلوتامین یا اوره تبدیل میکند. در برابر تامین نیتروژن، گیاه کربوهیدراتهای مورد نیاز (مالات) ریزوبیوم را تامین خواهد کرد. این باکتریها علاوه بر تثبیت نیتروژن در افزایش انحلالپذیری فسفر نقش دارند. سه نوع ساختار نودولی متفاوت در حبوبات وجود دارد. در تمام این نودولها آنزیم نیتروژناز کاملا از اثر اکسیژن محافظت میشود.
- نودولهای ثابت: در این نودولها اندکی پس از تشکیل ساختار، فعالیت مریستمها و تقسیم سلولی مهار میشود. در نتیجه سلولها تنها از راه افزایش حجم و طویل شدن رشد میکنند. نودولهای ثابت شکل کروی دارند. نودولهای ایجاد شده در ریشه دانه سویا از این نوع هستند.
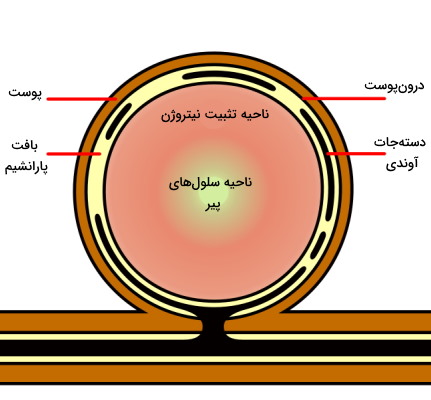
ریزوبیومها باکتریهای مهم در تثبیت نیتروژن و تغذیه گیاهی هستند که به شکل همزیست در نودولهای ریشه خانواده حبوبات زندگی میکنند. - نودولهای متغیر: نودولهای بیشتر حبوبات از این نوع هستند. علت نامگذاری این نودولها به دلیل حفظ سلولهای مریستمی راسی است که با تقسیم میتوز سلولهای جدید تشکیل میدهند و به رشد ساختار کمک میکنند. به همین دلیل ساختار این نودولها برخلاف انواع ثابت استوانهای است. چهار منطقه متفاوت در این نودولها وجود دارد.
- ناحیه I یا ناحیه مریستم فعال: سلولهای مریستمی در این منطقه وجود دارند که از تقسیم و تمایز آنها سلولهای نواحی دیگر ایجاد میشود.
- ناحیه II یا ناحیه آلودگی: سلولهای این ناحیه بزرگتر از ناحیه اول هستند و تقسیم سلولی آنها مهار شده است. باکتریها در این ناحیه وارد سلولهایی میشوند که اندامک آمیلوپلاست (اندامک ذخیره نشاسته) در سیتوپلاسم آنها وجود دارد.
- ناحیه III یا ناحیه تثبیت نیتروژن: سیتوپلاسم سلولهای این ناحیه از واکوئلهای بزرگ و باکتریوئیدهای (آمیلوبلاست حاوی باکتری) بسیاری تشکیل شده است که وظیفه تثبیت نیتروژن را بر عهده دارند. لگموگلوبین، پروتئینی مونومری با ساختاری شبیه به میوگلوبین ماهیچه و دارای گروه هم در سلولهای این ناحیه است که تمایل زیادی برای اتصال به مولکول اکسیژن دارد. این پروتئین با تنظیم غلظت اکسیژن آزاد در سیتوپلاسم، امکان فعالیت آنزیم دهیدروژناز و تنفس سلولی را فراهم میکند. این بخش نودول به دلیل وجود لگموگلوبین قرمز دیده میشود.
- ناحیه IV یا ناحیه سلولهای پیر: این قسمت از سلولها و باکتروئیدهای تجزیه شده تشکیل شده است. رنگ سبز انتهای این نودلها به دلیل تجزیه لگموگلوبین و آزاد شدن گروه هم آن ایجاد میشود.
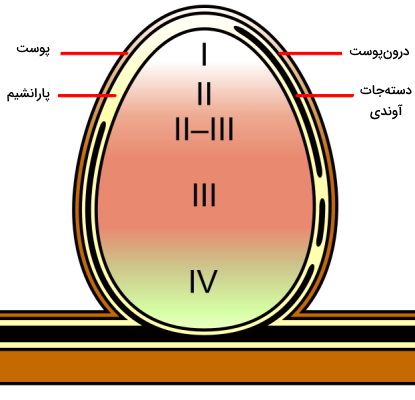
سلولهای نواحی مختلف نودولهای متغیر در مراحل متفاوت رشد و متابولیسم هستند.
- نودولهای اکتینوریزال: این نودولها ساختارهای متفاوت در گیاهان غیر حبوبات هستند. سلولهای مشتق از پوست ناحیه آلودگی را تشکیل میدهند.


سوالات متداول
در این بخش به تعدادی از سوالات متداول پیرامون تغذیه گیاهی پاسخ میدهیم.
کمبود کدام عناصر در تغذیه گیاهی بیشتر به وجود میآید ؟
فسفر، نیتروژن و آهن سه عنصری هستند که کمبود آنها در تغذیه گیاهی از خاک بیشتر ایجاد میشود. درصد فسفر موجود در خاک معمولا بسیار کمتر از نیاز گیاه است. نیتروژن عنصری حیاتی برای تغذیه گیاهی است که درصد ساختارهایی که گیاه توانایی جذب آن را ندارد در خاک بیشتر است. جذب آهن در خاک قلیایی برای برخی گیاهان مشکل است. برای حل این مشکل میتوان pH خاک بهوسیله اضافه کردن ترکیبات کلاتهکننده آهن کاهش داد.
مکانیسم های جذب مواد معدنی از خاک کدامند ؟
ریشه اندام اصلی برای جذب مواد معدنی و آب از خاک است. تار کشنده ساختاری باریک و بلند شبیه به تار مو و سلولهای مریستمی در راس آن محل اصلی ورود یونهای معدنی، عناصر معدنی و آب به گیاه است. انتشار ساده، اسمز و انتقال فعال سه مکانیسم اصلی برای ورود این مواد به ریشه و حرکت آنها در عرض ریشه تا آوند چوبی هستند.
جمع بندی
در این مطلب توضیح دادیم تغذیه گیاهی شامل دو دسته مواد معدنی و مواد آلی میشود. همچنین یاد گرفتیم ریشه وظیفه اصلی جذب مواد معدنی برای تغذیه گیاه را بر عهده دارد. ورود این عناصر به سلولهای ریشه از مسیرهای انتقال فعال (با مصرف انرژی) و انتشار ساده (بر اساس اختلاف غلظت الکتروشیمیایی) انجام میشود. جذب یونهای مختلف از ریشه یکسان نیست و عومال مختلفی ازجمله غلظت و نوع یونهای خاک، برهمکنش یونها با هم، دمای و pH خاک بر جذب عناصر و تغذیه معدنی گیاه اثر میگذارد. به علاوه از کودهای شیمیایی، آلی و زیستی در فواصل زمانی و مراحل مختلف رشد گیاهان برای بهینهسازی ترکیب خاک استفاده میشود.









