



تکوین چیست؟ | تعریف، انواع و مراحل | به زبان ساده
مطالعه تکوین جانوری در بسیاری از شاخههای علم مثل فیزیولوژی، پرستاری، پزشکی، زیستشناسی و حتی علوم شناختی و رفتاری اهمیت دارد. در این مقاله ضمن تعریف تکوین، در مورد انواع و روند آن در جانوران مختلف توضیح دادهایم.


تعاریف مورد استفاده
در این قسمت برخی عبارات و کلمات تخصصی که در متن به آنها اشاره شده است را توضیح دادهایم:
- «آمفیوکسوس» (Amphioxus): یک موجود دریایی از از نوع طنابداران است که گونهای بین مهرهداران و بیمهرگان محسوب میشود.
- «خارپوستان» (Echinoderms): موجودات آبزی مانند ستارهدریایی که بدن آنها عموما با خارهای آهکی پوشیده میشود.
- «ارکانترون» (Archenteron): حفره گوارش دیرینه، اولین حفره غذایی است که در تکوین و طی مرحله گاسترولا، در رویان ایجاد میشود.
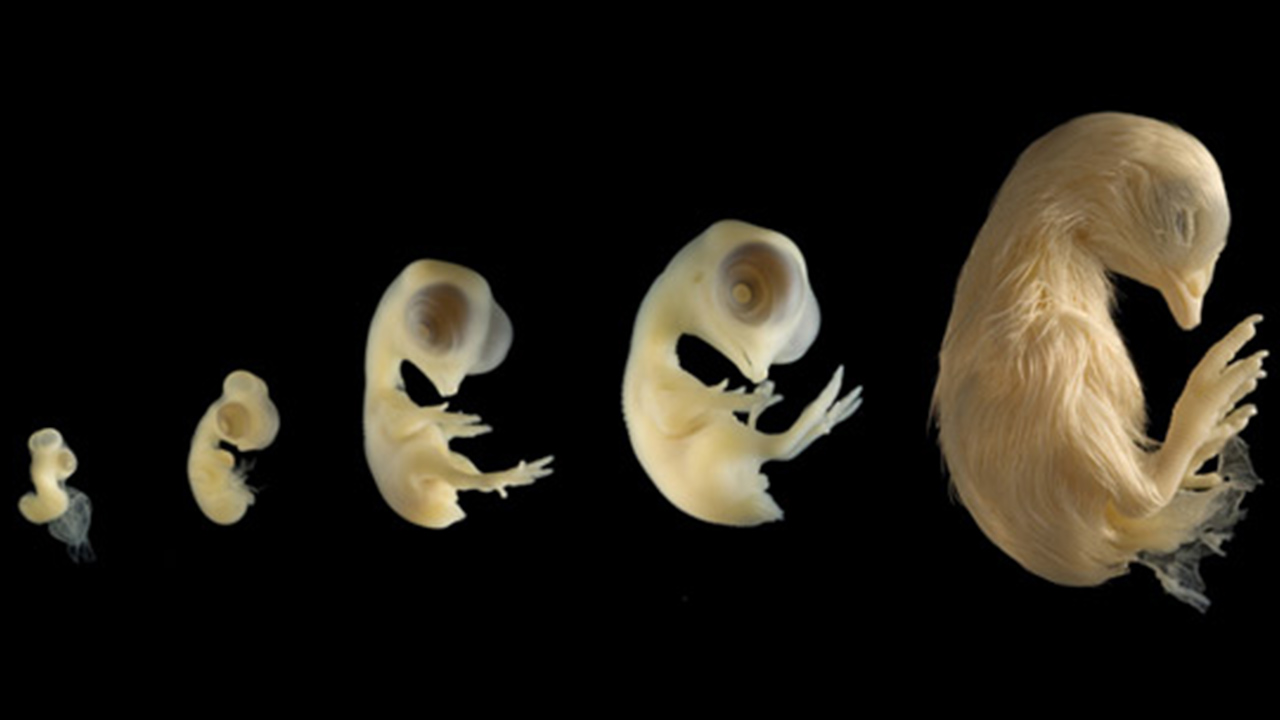
تکوین حیوانی چیست؟
فرایندی که طی آن، از سلول یک والد یا سلولهای دو والد، یک موجود زنده جدید به وجود میآید. در واقع تکوین طی تولید نسل جدیدی از ارگانیسمها شکل میگیرد.

تولید مثل و تکوین
در حیوانات پرسلولی (متازوآ)، «تولیدمثل» (Reproduction) به دو فرم جنسی و غیرجنسی رخ میدهد:
- «تولیدمثل جنسی» (Sexual Reproduction): جانور جدید نه از سلولهای سوماتیک یک والد، بلکه از سلولهای جنسی یا گامتهای دو والد به وجود میآید.
- «تولیدمثل غیر جنسی» (Asexual Reproduction): در این نوع تولید مثل، جانور جدید از سلولهای سوماتیک به وجود میآید. موجود جدید به صورت یک جوانه، از بلاستوما (گروهی از سلولهای والد) مشتق میشود. در برخی دیگر از گونهها همچون کرمها، ممکن است والد با تقسیم شدن خود به دو قسمت، تولیدمثل کند.
تولید مثل جنسی طی دورههای مختلف حیات در کره زمین، باعث ایجاد گونههای متنوع از جانداران و تکامل شده است.
مراحل ابتدایی تکوین
در جانوران پرسلولی، دو نوع سلول جنسی در تکوین موجود جدید، دخیل هستند:

هریک از این سلولها نیمی از ماده ژنتیکی لازم برای تولید مثل را فراهم میکنند و با یکدیگر ممزوج میشوند که فرایند لقاح گفته میشود. حاصل لقاح گامتهای نر و ماده، سلول تخم یا «زیگوت» (Zygote) است. محتوای سیتوپلاسمی تخم را گامت ماده فراهم و رویان طی رشد و تکوین اولیه خود از آن استفاده میکند. این منبع تغذیه در تخمگذاران، «زرده» (Yolk) نام دارد. زرده حاوی مواد مختلفی از جمله پروتئینها، فسفوپروتئینها، لیپیدها، کلسترول و چربی است که نوع، مقدار و نسبت آنها میتواند در گونههای مختلف، متفاوت باشد.
علاوه بر فراهم کردن مواد مورد نیاز جهت رشد و تکوین، سلول تخم باید برای تکوین رویان و پیدایش اندامهای مختلف، ساختار ویژهای در آن ایجاد میشود. برای شکلگیری رویان، تخم فرایندی به نام قطبزایی را طی میکند که حاصل آن ایجاد دو ناحیه در دو انتهای تخم است:
- «قطب جانوری» (Animal Pole): در این قطب هسته سلولی قرار گرفته و سیتوپلاسم فعالتر است. همچنین تقسیم میوز در قطب جانوری انجام میشود.
- «قطب نباتی» (Vegetal Pole) یا قطب گیاهی: سیتوپلاسم حضور چندانی در این ناحیه ندارد، اما حاوی مقدار بیشتری زرده است.
سازمانبندی حیوانی که تولید خواهد شد، با قطبیت سلول تخم ارتباط دارد.

انواع سلول تخم
در برخی از جانوران مانند بیمهرگان دریایی و پستانداران که جنین از خون مادر تغذیه میکند، تخم آنقدر کوچک است که معمولا با چشم غیر مسلح قابل مشاهده نیست. مثلا شعاع زیگوت در انسان به حدود ۰/۱ میلیمتر میرسد. انواع تخمها بر اساس میزان ماده زرده طبقهبندی میشوند:
- «تخم اولیگولسیتال» (Oligolecithal Egg): میزان زرده ذخیره شده در تخم نسبتا کم و توزیع آن یکنواخت است.
- «تخم تلولسیتال» (Telolecithal Egg): زرده بیشتری دارد که به صورت غیرِ یکنواخت در تخم توزیع میشود. به طور مثال ممکن است در قطب نباتی مقدار بیشتری زرده وجود داشته باشد.
- «تخم مگالسیتال» (Megalecithal Egg): بیشترین مقدار زرده در این نوع تخم وجود دارد که درون تعدادی گرانول یا یه صورت یک توده فشرده، داخل تخم قرار گرفته است.

تکوین چه مراحلی دارد؟
درک روند تکوین با تقسیمبندی آن به مراحل مختلف آسانتر است. در هر مرحله تغییراتی به وجود میآیند تا سلول تخم به هدف خود یعنی تشکیل یک موجود زنده سازمانیافته برسد. بر اساس نیازهای لازم برای انتقال از هر مرحله به مراحل بعدی، میتوان تکوین را به بخشهای مختلفی تقسیمبندی کرد که در ادامه آنها را توضیح دادهایم.
کلیواژ یا تسهیم چیست؟
بلافاصله پس از لقاح و تشکیل تخم، «کلیواژ» (Cleavage) یا تسهیم رخ میدهد. کلیواژ با سایر انواع تقسیم سلولی متفاوت است زیرا طی آن ماده ژنتیکی و تعداد یاختهها افزایش مییابند اما مقدار ماده سیتوپلاسمی ثابت میماند. از آنجایی که هدف از تکوین، تولید یک ارگانیسم پرسلولی است، باید از زیگوت تکسلولی، تعداد زیادی سلول جدید به وجود بیاید. این فرایند طی کلیواژ یا تسهیم انجام میشود که مجموعهای از تقسیمهای سلولی متوالی از نوع میتوز است. سلولهایی که طی کلیواژ به وجود میآیند «بلاستومر» (Blastomeres) نام دارند.
طی کلیواژ و در فاصله بین تقسیمات متوالی، تقریبا هیچ رشد سلولی صورت نمیگیرد و حجم کلی ماده سلولی مقدار ثابتی دارد. در نتیجه اندازه سلولها طی هر تقسیم، تقریبا به نصف کاهش مییابد. در آغاز روند کلیواژ، تقسیمات سلولی در تمام بلاستومرها همزمان صورت میگیرند و تعداد سلولها در هر دور تقسیم دو برابر میشود اما با جلو رفتن این روند، سلولها دیگر به صورت همزمان تقسیم نمیشوند.

انواع کلیواژ جانوری
کلیواژ سلولهای متفاوتی ایجاد میکند که بلاستومر نام دارند و یک توده فشرده به نام مورولا را به وجود میآورند. در نهایت کلیواژ باعث ایجاد بلاستولا خواهد شد. بر اساس میزان زرده موجود در تخم، کلیواژ به انواع مختلفی انجام میشود.
کلیواژ محدود
«کلیواژ محدود» (Determinate) یا کلیواژ موزاییکی، در بسیاری از «دهاننخستیان» (Protostome) انجام میشود. حاصل این نوع کلیواژ بلاستومرهایی هستند که برخی قدرت تبدیل به رویان را ندارند.
کلیواژ نامشخص
در این نوع از تسهیم بلاستومرهای به وجود آمده، ویژگیهای سلولی نباتی/جانوری دستنخوردهای دارند و هریک از آنها میتوانند یک ارگانیسم کامل را ایجاد کنند. «کلیواژ نامشخص» (Indeterminate) در «دوتروستومها» (Deuterostomes) دیده میشود.
کلیواژ هولوبلاستیک
اگر زرده موجود در تخم بارور کم باشد، چهار نوع کلیواژ امکانپذیر هستند که همگی زیر شاخه کلیواژ هولوبلاستیک محسوب میشوند. تقسیم اول در این نوع تسهیم همیشه در راستای محور جانوری/نباتی و دومین کلیواژ عمود بر تقسیم اول است. انواع کلیواژ هولوبلاستیک عبارتند از:
- «دو طرفه» (bilateral): دو سلول به وجود آمده کاملا از یکدیگر جدا میشوند.
- «شعاعی» (radial): در برخی ستارههای دریایی و مهرهداران وجود دارد که در آن محور دوکی موازی یا عمود بر محور قطبی اووسیت است.
- «چرخشی» (rotational): تفاوت این نوع کلیواژ در تقسیم دوم است، یکی از سلولها بر محور افقی و دیگری بر محور استوایی تقسیم میشوند. در برخی پستانداران که مقدار خیلی کمی زرده وجود دارد این نوع کلیواژ دیده میشود و تخم به فاصله کوتاهی از تقسیم باید در دیواره رحم جایگزین شود تا بتواند تغذیه کند. یکی از مشهورترین جانداران با این مدل از کلیواژ، C.الگانس است.
- «مارپیچی» (spiral): تسهیم مارپیچی به دو حالت تقسیم برابر و نابرابر انجام میگیرد. در نتیجه این نوع کلیواژ چهار نوع بلاستومر A، B، C و D ایجاد میشوند. این نوع کلیواژ در جانورانی مانند کرمهای حلقوی، برخی کرمهای روبانی و نواری وجود دارد.
کلیواژ مروبلاستیک
اگر مقدار زیادی زرده در تخم بارور شده وجود داشته باشد، تخم میتواند وارد کلیواژ نسبی یا مروبلاستیک شود که خود دو نوع دارد:
- «دیسکی» (Discoidal): دیسک تقسیم از زرده عبور نمیکند. در پرندگان، خزندگان و ماهیهایی با تخم تلولسیتال دیده میشود.
- «سطحی» (Superficial): میتوز بدون تقسیم سیتوپلاسم اتفاق میافتد و در نتیجه سلول چند هستهای ایجاد میشود. این نوع کلیواژ در بندپایان مشاهده میشود.

کلیواژ در جانوران
در بیشتر جانوران کلیواژ یک الگوی منظم دارد. اولین تقسیم، درون صفحه محوری زیگوت رخ میدهد. صفحه تقسیم، با این فرض که محور اصلی تخم عمودی است، به صورت قراردادی عمودی در نظر گرفته میشود. دومین صفحه تقسیم سلولی نیز عمودی اما با زاویه ۹۰ درجه نسبت به صفحه تقسیم قبلی است و بنابراین باعث ایجاد یک ضلع جدید در توده سلولی میشود. سومین صفحه کلیواژ هم با زاویه ۹۰ درجه نسبت به هر دو صفحه قبلی و وضعیتی افقی یا استوایی دارد.
تقسیمات بعدی ممکن است به تناوب بین صفحات کلیواژی عمودی و افقی انجام شوند اما جهتگیری تقسیمات سلولی در مراحل آخر کلیواژ تصادفی هستند. گرچه این الگو در بسیاری از جانوران مشابه است اما الگوهای پیچیدهتری در برخی حیوانات مانند کرمهای حلقوی، نماتودها و حلزونها دیده میشود. هرچه میزان زرده در تخم بالاتر باشد، با ایجاد مانع در مسیر تقسیم سلولی، بر روند کلیواژ اثر میگذارد.
در بلاستومرهای تخمهای اولیگولسیتال، مقدار زرده کمتر بوده و درون وزیکولهایی قرار دارد که از حرکات سیتوپلاسمی تبعیت میکنند. اما در تخمهای مگالسیتال با زرده فراوان، کلیواژ در نزدیکی قطب جانوری انجام میشود که زرده کمتری در آن قسمتها وجود دارد. در نتیجه بلاستومرهای نزدیک به قطب جانوری کوچکتر از بلاستومرهای نباتی هستند.
تراکم بالای زرده ممکن است باعث به تعویق افتادن کلیواژٰ در یک یا تمام قسمتهای زیگوت شود که در این صورت کلیواژ فقط در قطب جانوری رخ میدهد و کلیواژ نوع «مروبلاستیک» (Meroblastic) نام دارد. در این صورت فقط قسمتی از ماده موجود در زیگوت، بین سلولها تقسیم میشود و باقی آن به عنوان منبع غذایی برای رشد و نمو جنین، متراکم باقی میماند.
کلیواژ در بی مهرگان
کلیواژ در بسیاری از بی مهرگان مانند کیسهتنان، کرمهای حلقوی، خارپوستان، طنابداران و سرطنابداران، از نوع کامل یا «هولوبلاستیک» (Holoblastic) است. اندازه بلاستومرها ممکن است تقریبا مساوی یا فقط کمی متفاوت باشند. کلیواژ در دوزیستان از نوع هولوبلاستیک است اما اندازه بلاستومرها بسیار نابرابر هستند. بلاستومرها در قطب جانوری کوچکترین و در قطب نباتی بزرگترین اندازه را دارند. در بسیاری از حلزونیان هم به همین شکل است.
در بسیاری از ماهیها، پرندگان، خزندگان و پستانداران تخمگذار، کلیواژ دیسکی است یعنی به یک دیسک سیتوپلاسمی در قطب جانوری محدود است و بیشتر ماده زردهای تقسیم نشده باقی میماند. کلیواژ در حشرات و دیگر بندپایان سطحی است به این معنی که، کل لایه سطحی سیتوپلاسم تخم، به سلولهای جدیدی تقسیم میشود و تخم دارای یکی زرده تقسیم نشده و متراکم مرکزی خواهد بود. شرایط کلیواژ در پستانداران جفتدار مانند انسان، ویژه است.
وقایع سلولی در کلیواژ
طی کلیواژ، تکوین فقط شامل افزایش تعداد سلولها است، شکل رویان تغییر نمیکند و تحولات شیمیایی آن به تغییراتی که برای تقسیم سلولی ضروری هستند، محدود میشود. تحولات شیمیایی و ساختاری با تجمع ماده کروموزومی در هسته بلاستومرها مرتبط هستند. قبل از هر تقسیم، کروموزومها حاوی دو دسته ژن هستند و این یعنی ماده کروموزومی (DNA) پیش از تقسیم همانندسازی شده است. برای همانندسازی DNA نیاز به RNA، عناصر معدنی و مولکولهای آلی مورد نیاز هستند.
در صورت تیتراسیون تخم یا پورومایسین (مادهای که سنتز پروتئینها را مهار میکند) کلیواژ سریعا متوقف میشود که نشان میدهد وجود مقدار مشخصی پروتئین نیز برای پیشبرد کلیواژ ضروری است. نوع پروتئینهای مسئول در این فرایند هنوز به خوبی شناخته نشدهاند اما بر اساس وجود هیچ پروتئینی نمیتوان تمایز بخشهای مختلف رویان را پیشبینی کرد. تصور بر این است که ژنهای کروموزومی در خلال کلیواژ غیرفعال باقی میمانند. سرعت کلیواژ کاملا به سیتوپلاسم تخم بستگی دارد.
گرچه شکل و حجم رویان طی کلیواژ تغییر نمیکند اما تغییرات مهمی در سازماندهی سلولها اتفاق میافتد. با تولید و حرکت بلاستومرها به طرفین، یک حفره پر از مایع در مرکز باقی میماند. در تسهیم هولوبلاستیک، بلاستومرها به صورت یک لایه با ضخامت یک تا چند سلولی، در اطراف این حفره قرار دارند.
رویان در این مرحله مانند یک توپ توخالی به نظر میرسد که «بلاستولا» (Blastula) گفته میشود. لایه خارجی سلولها «بلاستودرم» (Blastoderm) و حفره پر از مایع «بلاستوسل» (Blastocoel) نام دارند. در کلیواژ دیسکی، سلولها کل رویان را در بر نمیگیرند , تنها در قطب جانوری قرار دارند، در عین حال بلاستوسل ممکن است با یک شکاف بین بلاستومرها و توده زردهای به وجود بیاید. بلاستومرها سپس به شکل یک بلاستودیسک بشقاب مانند، بلاستوسل را میپوشانند.
تشکیل بلاستولا پایان مرحله کلیواژ را نشان میدهد. در مرحله بعدیِ تکوین، تغییر زیادی در تعداد سلولها ایجاد نمیشود و تقسیمات سلولی با سرعت کمتری ادامه خواهند یافت. این تقسیمات، نظم توده سلولیِ موجود را تغییر و به آن نظم میدهند تا به شکل نهایی حیوان تبدیل شود.

تکوین دستگاه گوارش
رویان در مرحله بلاستولا و قبل از سازمانیابی به سمت موجود بالغ، باید دچار تغییراتی شود. هر موجود پرسلولی بالغ در روند تکوین، اغلب بافتهایی با ساختار متحدالمرکز ایجاد میکند. این ویژگی در تمام جانوران پیشرفتهتر از اسفنج دریایی وجود دارد. بافتهای بالغ از رویان سه لایهای به نام «لایه زایا» یا زاینده (Germinal Layer) اشتقاق مییابند:
- «اکتودرم» (Ectoderm): لایه زایای خارجی که سطح پوششی پوست، سیستم عصبی و ارگانهای حسی را به وجود میآورد.
- «مزودرم» (Mesoderm): لایه زایای میانی، سلولهای عضلانی، اندامهای دفعی، اندامهای گردشی، گنادها (اندامهای جنسی) و اسکلت داخلی را ایجاد میکند.
- «اندرودرم» (Endoderm) یا «انتودرم» (Entoderm): داخلیترین لایه زایا، کانال گوارشی و بخشهای مرتبط با گوارش و تنفس را تشکیل میدهد.
بلاستولا که فقط دارای یک لایه سلولی است، با یک تغییر ساختاری در بلاستومرها، ارگانهای مختلف سیستم بدن موجود زنده را به وجود میآورد. این کار طی فرایند گاسترولاسیون انجام میشود که شامل تغییر یا حرکت ماده سلولی رویانی است تا سه لایه زایا در موقعیت صحیح خود قرار بگیرند.
تغییر جایگاه بلاستولا برای ایجاد لایههای زایا، به خوبی در انواع خاصی از جانوران دریایی با زیگوت اولیگولسیتال مشاهده میشود. بلاستولای توخالی، شامل یک لایه اپیتلیال ساده (بلاستودرم) است که تغییر آن را میتوان به کشیدن توپ پلاستیکی به یک سمت تشبیه کرد. در نتیجه چنین کششی، رویانِ کروی به شکل یک فنجان دو جداره در میآید و روزنه روی آن، محل قطب نباتی سابق را نشان میدهد.
قسمتی از بلاستودرم که جدار داخلی فنجان را پوشانده است و به اندودرم و مزودرم تبدیل میشود و بلاستومرهایی که در لایه خارجی باقی ماندهاند نیز اکتودرم را ایجاد میکنند. به دنبال چنین بی نظمی در قطب نباتی، بلاستوسل کاهش مییابد یا از بین میرود و یک حفره جدید تشکیل میشود که همان حفره رودهای اولیه یا آرکنترون است. این حفره در نهایت به یک هسته لومنی توخالی به نام «کانال گوارشی» (Alimentary Canal) تبدیل میشود. در این مرحله، رویان یک روده ابتدایی با یک روزنه خارجی به نام گاسترولا دارد. روزنه گاسترولا، «بلاستوپور» (Blastopore) یا دهانِ ابتدایی گفته میشود. عبارت دهان ابتدایی گمراهکننده است چون این روزنه همیشه در بالغین به دهان تبدیل نمیشود و در برخی گروههای خاص جانوری این روزنه مقعد را ایجاد میکند، بنابراین نام بلاستوپور صحیحتر به نظر میرسد.
در برخی از کیسهتنان، سلولهای قطب نباتی «پاکت تو رفته» (Invaginating Pocket) را ایجاد نمیکنند اما سلولهای خاصی به سمت داخل رفته و ارتباط خود را با سایر سلولهای بلاستودرم از دست میدهند. در نهایت این سلولها بلاستوسل را پر میکنند و یک توده متراکم اندودرمی را به وجود میآورند. حفره درون این توده متراکم (آرکنترون) و منفذ (بلاستوپور)، با جدا شدن این سلولها، به صورت ثانویه تولید میشوند.
در آموزش زیر به اجزا، ساختار و عملکرد دستگاه گوارش انسان به طور مفصل پرداختهایم و مطالعه آن را توصیه میکنیم.
- برای مشاهده آموزش دستگاه گوارش انسان، عملکرد و اجزای آن | به زبان ساده + اینجا کلیک کنید.

تکوین در آمفیوکسوس، سخت پوستان و دوزیستان
گاسترولاسیون همیشه طبق روندی که توضیح داده شد، انجام نمیشود. طی تکامل، در برخی گروههای جانوری خاص، این مراحل تغییراتی پیدا کردهاند که بدون شک در جهت بقای گونهها بوده است. به طور مثال، در آمفیوکسوس که یک طنابدار ابتدایی و شبه ماهی است، بلاستودرم تو رفته، نهایتا در تماس نزدیک با سطح داخلی اکتودرم قرار میگیرد در نتیجه عملا بلاستوسل را به سمت بیرون هل میدهد یا حداقل موجب کاهش اندازه آن تا حد یک شکاف باریک بین اکتودرم و اندومزودرم میشود.
در خارپوستان، بخش کوچکتری از بلاستودرمِ تورفته و بلاستوسل، به عنوان یک حفره داخلی ویژه بین اکتودرم و اندومزودرم باقی میماند که حفره بدنی ابتدایی و تنها حفره بدن (به جز حفره کانال گوارشی) در بیمهرگانی مانند نماتودها و روتیفرها خواهد بود.
در مرحله double-walled-cup، دو لایه زایای داخلی یعنی اندودرم و مزودرم، ممکن است هنوز متمایز نشده باشند و این تمایز بعدا در فاز دوم از گاسترولاسیون و از طریق یکی از دو روشی که در ادامه توضیح داده شده است، رخ بدهد.
- روش اول، تکوین کیسه خارجی از دیواره آرکنترون است. در ستارهماهیها و دیگر کیسهتنان بخش عمقی اندومزودرم تورفته، یک ساک با دیواره نازک در هر طرف گاسترولا ایجاد میکند که پیشسازهای مزودرم هستند، بخش باقیمانده آرکنترون، تبدیل به اندودرم میشود و دیواره روده را تولید میکند. حفره موجود در ساکهای مزودرمی برای تبدیل به حفره ثانویه یا «سلوم» (Coelom)، اتساع مییابد. روند مشابه با تکوین مزودرم و سلوم در آمفیوکسوس، در طنابداران هم رخ میدهد، با این تفاوت که تعدادی از ساکهای مزودرمی در هر دو طرف رویان تشکیل میشوند که بعدا ساختارهای قطعه قطعه رایج در طنابداران را به وجود میآورد. تنها قدامیترین جفت ساکهای مزودرمی، در زمان شکلگیری خود دارای حفره هستند و ساکهای خلفیتر، سلولهای متراکمی هستند که از دیواره archenteric و از یکدیگر جدا شدهاند و بعدتر، حفرههای سلومی را ایجاد خواهند کرد.
- روش دوم شکلگیری مزودرم به وسیله جدا شدن سلولهای مزودرم، از توده مزودرمی است. این رخداد ممکن است باعث ایجاد سلولهایی جدا از یکدیگر و جدا از حفره archenteric یا جدا از کل لایههای سلولی شکافته شده از اندودرم شود. در گاسترولاسیون دوزیستان این نوع فرایند دیده میشود.
تکوین نواحی خاص در رویان دوزیستان (که با رنگآمیزی طبیعی یا استفاده از رنگهای مصنوعی مشخص شدهاند) و محل آنها در دوزیست بالغ به صورت طرحی تحت عنوان نقشه سرنوشت قابل ردیابی هستند. نقشه سرنوشت بلاستولای قورباغه قبل از مرحله گاسترولاسیون، نشان میدهد که مواد لازم برای تشکیل ارگانهای مختلف رویان هنوز در محل نهایی خود در حیوان بالغ، دیده نمیشوند. به طور مثال، ماده اندودرمی لازم برای «روده قدامی» (Foregut)، فاصله زیادی با قطب نباتی ندارد، عناصر اکتودرمی ناحیه دهانی (Stomodeum) نیز در نزدیکی قطب جانوری قرار دارند. بنابراین بازآرایی وسیعی برای قرار گرفتن تمام این بخشها در محل هدف و ایجاد ارتباط عملکردی صحیح بین آنها نیاز است.
به دلیل وجود مقدار زیاد زرده و کلیواژ نامساوی، گاسترولاسیون در دوزیستان با تورفتگی ساده در قطب نباتی، قابل انجام نیست. مقدار مشخصی از تورفتگی، همراه با گسترش فعال نیمکره جانوریِ رویان رخ میدهد و در نتیجه اکتودرم، نواحی اندودرم و مزودرم را میپوشاند. گاهی از گسترش با عنوان رشد زیاد یاد میشود که یک عبارت نادرست است زیرا هیچ رشد یا افزایش تودهای اتفاق نمیافتد. اکتودرم در نهایت نازک میشود، گسترش مییابد و سطح بزرگتری از رویان را در نوعی از جابجایی سلولی به نام «اپیبولی» (Epiboly) میپوشاند. اپیبولی در مرحله گاسترولاسیون رخ میدهد.
گاسترولاسیون در دوزیستان، «ریهماهی» (Lungfishes) و «سیکلوستومها» (Cyclostomes) با تشکیل یک حفره روی قسمتی که بخش پشتی رویان را خواهد ساخت، آغاز میشود. این حفره، نشاندهنده مهاجرت سلولی به درون بلاستودرم است. در حالیکه این سلولها تغییر شکل پیدا میکنند، با کشیده شدن سلولهای مجاور به مرکز، حتی قبل از کاهش، ادغام در سطح خارجی نیز به وجود میآید. سلولهایی که بیشترین درگیری را در این فرایند دارند بخشی از روده قدامی را تشکیل خواهند داد. حرکات سلولی بعدی به درون، یک حفره تشکیل میدهد که به سرعت به یک آرکنترون پاکت مانند و بلاستوپور تبدیل میشود.
پس از تشکیل آرکنترون، تعداد بیشتری از سلولهای خارجی از لبه بلاستوپور به داخل میروند. در دوره گاسترولاسیون، شکل بلاستوپور از یک حفره ساده به یک شکاف عرضی و نهایتا به یک گودی که در قطب نباتی به دور زرده حلقه زده تغییر میکند.
یکی از نتایج اپیبولی همیسفر جانوران این است که لبه بالایی شکاف، کم کم به پایین کشیده میشود تا سلولهای زرده در قطب نباتی، به طور کامل پوشیده شوند. سپس حاشیه بلاستوپور به سمت قطب نباتی همگرایی پیدا میکنند و شیار بین آنها به یک کانال در انتهای خلفی رویان تقلیلی پیدا میکند. این کانال در برخی گونهها، سوراخ آنالی (مقعدی) را به وجود میآورد (در سایر موارد، کانال بسته میشود و یک منفذ مقعدی در مجاورت آن به وجود میآید که در ناحیه شکمیتری قرار دارد).
حفره آرکنترون با انتقال بیشتر ماده از خارج به داخل، بزرگتر میشود. مزودرم و اندودرم به داخل منتقل میشوند و فقط اکتودرم در سطح رویان باقی میماند. مزودرم از اندودرم جدا میشود: اندودرم در راستای حفره آرکنتریکی قرار میگیرد (و نهایتا کانال غذایی را میسازد). همزمان مزودرم، اندودرم را میپوشاند تا «مزودرم محوری» (Axial Mesoderm) یا «Chordamesodermal Mantle» را به وجود بیاورد. هنگام بسته شدن بلاستوپور، سه لایه زاینده ارتباط فضایی درستی با یکدیگر تشکیل دادهاند.

تکوین در خزندگان، پرندگان و پستانداران
گرچه گاسترولاسیون دوزیستان در مقایسه با جانورانی با تخم الیگولسیتال تغییر یافته است، آرکنترون طی فرایند تورفتگی به وجود میآید. در هر حال در مهرهداران پیشرفته، که تخمهایی با مقدار فراوان زرده تولید میکنند مانند خزندگان، پرندگان و پستانداران تخمگذار، چنین روندی رخ نمیدهد. کلیواژ در این حیوانات نسبی یا «مروبلاستیک» (Meroblastic) است. در نهایت رویان شامل یک گروه از سلولها با نظم دیسکی شکل خواهد بود که در بالای توده زردهای قرار گرفتهاند.
این گروه سلولی اغلب به یک لایه بالایی به نام «اپیبلاست» (Epiblast) و لایه پایینی به نام «هیپوبلاست» (Hypoblast) تقسیم میشود. این لایهها معادل اکتودرم و اندودرم نیستند، چون تقریبا تمام سلولهای ایجاد کننده رویان، در اپیبلاست حضور دارند. سلولهای مزودرمی و اندودرمی آینده، به داخل کشیده میشوند و تنها ماده اکتودرمی در سطح باقی میماند. در خزندگان، پستانداران تخمگذار و برخی پرندگان، یک کاهش پاکت مانند در اپیبلاست ایجاد میشود که فقط کوردامزودرم یا حتی فقط «پُشتمازه» یا «طناب پشتی» یا «نوتوکورد» (Notochord) را احاطه میکند.
معدود سلولهای باقیمانده از مزودرم و اندودرم به داخل مهاجرت میکنند و در آنجا به یک لایه کوردامزودرم و اندودرم سازماندهی میشوند. اگر چنینی لایه ای وجود داشته باشد، سلول دوم با سلولهای هیپوبلاست مخلوط میشود. مهاجرت سلولها با هدف ایجاد مزودرم و اندودرم در تمام سطح رویان دیسکی شکل انجام نمیشود و محدود به یک ناحیه خاص در خط میانی آن است. این ناحیه در خزندگان و به میزان کمتری در پستانداران رده پایین، تقریبا بیضی شکل است و در پستانداران رده بالا و پرندگان «خط ابتدایی» (Primitive Streak) نام دارد. قسمتی از اپیبلاست که متراکم و در انتهای قدامی ضخیمتر است، «گره هنسن» (Hensen’s Node) گفته میشود.
در حیواناتی که کلیواژ دیسکی دارند، سه لایه سلولی زاینده در انتهای گاسترولاسیون روی هم قرار میگیرند به طوری که اکتودرم در بالا، مزودرم در وسط و اندودرم در پایین قرار بگیرند. رویان از لایههای مسطحی تشکیل شده است که با فرایند تورفتگی یک سیستم از لایههای متحدالمرکز را بسازد. حاشیه لایههای زایا که در فرایند تورفتگی نقشی ندارند، به زرده متصل باقی میمانند و بخشهای خارج رویانی را تشکیل میدهند و مستقیما در فراهم کردن سلولها برای رویان درگیر نیستند اما زرده را تقسیم میکنند و به رویان در حال تکوین انتقال میدهند.
پستانداران پیشرفته - در تقابل با پستانداران تخم گذار- در زیگوت خود زرده ندارند اما در حیواناتی که تخم آنها زرده دارد، بعد از عبور از یک مرحله تکاملی، خصوصا در مرحله گاسترولاسیون، زرده باقی میماند که این ویژگی در خزندگان رایج است (و در پرندگان که نیای مشترک با خزندگان دارند). در نتیجه، در انتهای کلیواژ، سلولهای قبلی رویان (سلولهایی که در نهایت بدن حیوان را میسازند) به شکل یک دیسک اطراف یک حفره آرایش مییابند که در زرده خزندگانی با نیای مشترک با پستانداران اتفاق میافتد. در این دیسک سلولی یک ورقه ایجاد میشود که تعداد لایههای زاینده آن با آنچه در پرندگان و خزندگان دیده میشود مشابه است.
گاسترولاسیون و تشکیل سه لایه زاینده، آغاز تقسیم توده سلولی رویانی و کلیواژ است. سپس سلولها تحت تاثیر مستقیم ژنها تغییر و تحول مییابند. ژنهایی که از اسپرم آمدهاند برای اولین بار وقایع سلولی را کنترل میکنند چون به نظر میرسد که همه وقایع کلیواژ، با ژنهای مادری کنترل میشوند. در هیبریدیزاسیون که طی آن از هر موجود، گونههای مختلفی ایجاد خواهند شد، اثر اسپرمی اولین مشاهده در گاسترولاسیون است: ویژگیهای پترنال ممکن است در این مرحله مشاهده شوند. یا اگر ژنهای پدری با تخم ناسازگار باشند، ممکن است تکوین رویان متوقف شود و از بین برود (مثل هیبرید شدن گونههایی که خویشاوندی ندارند).
تنوع سلولها در رویان به سرعت، در خلال و بعد از گاسترولاسیون افزایش مییابد. در نتیجه، لایه های زاینده به سرعت به تعداد زیادی سلولها تقسیم میشوند که شکل ابتدایی و اولیه ارگان و اندامهای مختلف و سیستمیک رویان هستند و پس از گاسترولاسیون تولید اندام یا اندامزایی روی خواهد داد.










با سلام و تشکر از مطالب بسیار مفید و ارزنده شما لطفاً در خصوص
“زیست فناوری و ارتباط آن با علوم شناختی” اگه مطلبی دارید برام بفرستید، ممنون میشم.
عاااالى بود
کلیات آن خیلی عالی بود ممنون ولی به نظرم جزیات آن کمی زیاد بود فکر رامشغول میکرد بازهم سپاس
خیلی کامل و مفید👍