



یاخته های بافت عصبی – انواع و عملکرد به زبان ساده
بدن جانوران از بافتهای مختلفی تشکیل شده است که هر کدام عملکرد و ساختار متفاوتی دارند. برای مثال بافت عصبی به جاندار کمک میکند محیط اطراف خود را درک کند و با ایجاد پاسخ مناسب به محرکهای محیطی در حفظ بقا و برقراری ارتباط موجود با دیگر افراد هم گونه و محیط نقش فراوانی دارد. در این مطلب انواع یاخته های بافت عصبی و نقش هر کدام در این بافت را توضیح میدهیم.
- خواهید آموخت که نورونها و یاختههای پشتیبان چگونه با هم همکاری میکنند.
- یاد میگیرید انواع انتقالدهنده عصبی و نقش هر کدام را تشخیص دهید.
- میآموزید سازوکار انتقال پیام عصبی و عملکرد سیناپس را تحلیل کنید.
- خواهید توانست نقش اختلال عملکرد سلولهای گلیال را در بیماریهای عصبی توضیح دهید.
- پتانسیل عمل و نقش کانالهای یونی را در انتقال عصبی بررسی میکنید.
- درک جامعی از تفاوتهای عملکردی بین سیستم عصبی مرکزی و محیطی کسب میکنید.
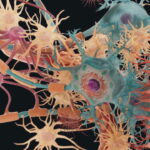
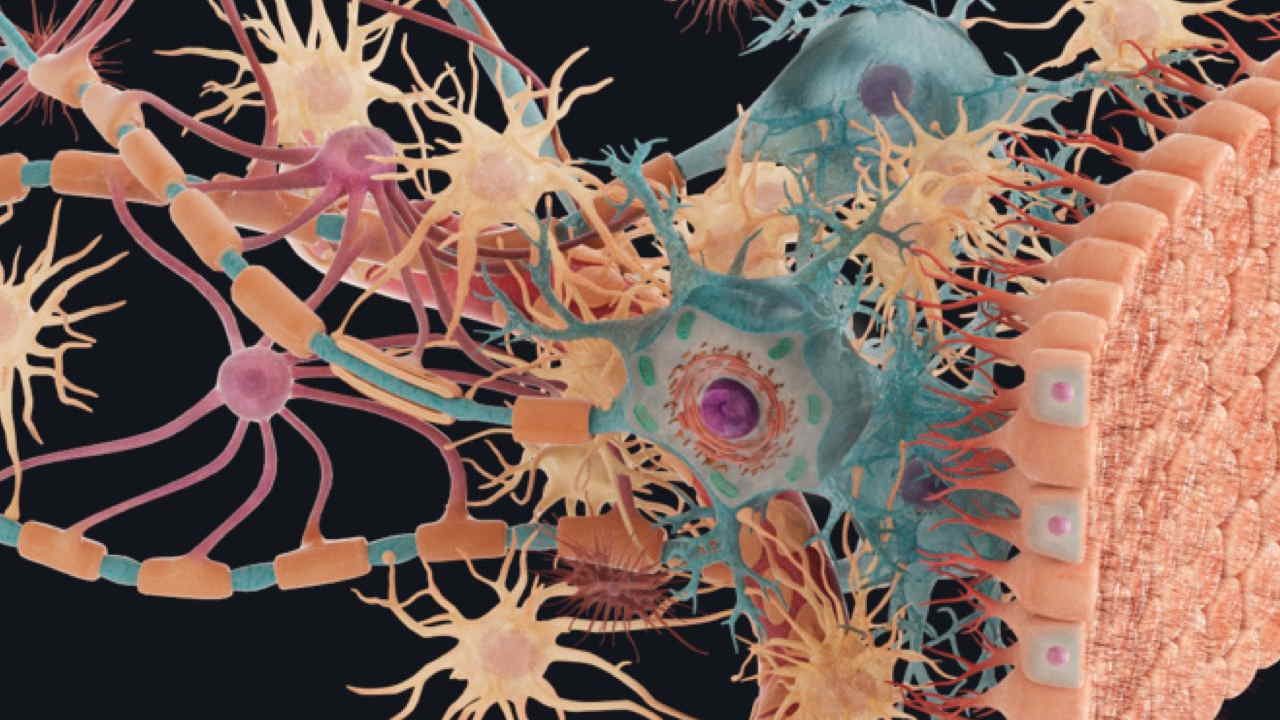


بافت عصبی چیست ؟
بافت عصبی شبکهای از سلولهای هدایتکننده پیام الکتریکی در بدن مهرهداران است که در اکثر اندامهای جانور وجود دارد و عملکردهای مختلف آن ازجمله درک محرکهای محیطی، حرکت و تنظیم هومئوستازی را بر عهده دارد. این بافت به دو بخش «سیستم عصبی مرکزی» (Central Nervous System | CNS) و «سیستم عصبی محیطی» (Peripheral Nervous System | PNS) تقسیم میشود.
- سیستم عصبی مرکزی: این سیستم از مغز و نخاع تشکیل شده است. یاختههای عصبی این بخش پیام عصبی بخشهای مختلف بدن را دریافت و با تحلیل پیام امکان پاسخ مناسب به محیط را فراهم میکند.
- سیستم عصبی محیطی: این بافت عصبی بین اندامهای مختلف و سیستم عصبی مرکزی ارتباط برقرار میکند. برای مثال در حرکت بازو پیام از سیستم عصبی مرکزی به محیطی و سپس ماهیچه منتقل میشود. این سیستم از رشتههای عصبی سمپاتیک، پاراسمپاتیک و سیستم حسی تشکیل شده است.
- اعصاب سمپاتیک: این رشتههای عصبی مکانسیمهای فرار یا حمله (Fight-or-Flight) را تنظیم میکنند. در مواقع ایجاد خطر برای بافت یا اندام این اعصاب فعال میشود. تحریک اعصاب سمپاتیک باز شدن مردمک چشم، افزایش ضربان قلب افزایش تعریق و مهار گوارش را به همراه دارد.
- اعصاب پاراسمپاتیک: این رشتههای عصبی در زمان استراحت و گوارش (Rest and Digest) فعال میشوند. تحریک این اعصاب گوارش، دفع و برانگیختگی جنسی را تنظیم میکند و ضربان قلب را کاهش میدهد.
- اندامهای حسی: گیرندههای حسی انتهای آکسونی تغییرشکلیافته هستند که پیامهای شیمیای، تغییر فشار، تغییر شدت نور و تغییر دما را به جریان الکتریکی تبدیل میکنند. رشتههای اعصاب محیطی این پیام را برای تحلیل و پاسخ به سیستم عصب مرکزی انتقال میدهند.
انواع یاخته های بافت عصبی کدامند ؟
بافت عصبی سیستم بسیار پیچیدهای است که برای برقراری ارتباط و تنظیم فعالیت اندامها و بافتهای مختلف بدن از میلیونها سلول تشکیل شده است. نورونها یاخته های اصلی بافت عصبی هستند که پیامها را به شکل جریان الکتریکی از اندامها به مغز و نخاع انتقال میدهند. اما این سلولها برای عملکرد خود نیاز به یاخته های پشتیبان «نوروگلیا یا گلیا» (Neuroglia or Glial) دارند.
یاخته های پشتیبان
یاخته های پشتیبان یا سلولهای گلیا، سلولهای غیرعصبی هستند که در سیستم عصبی مرکزی و محیطی در کنار نورونها قرار دارند. این سلولها در برابر آسیبهای فیزیکی از نورون محافظت و متابولیسم این سلولها را پشتیبانی میکنند. میتوان گفت یاخته های پشتیبان بافت پیوندی ویژه در سیستم عصبی هستند. ساختار و عملکرد این سلولها با نورونها متفاوت است. یاخته های پشتیبان سیستم عصبی مرکزی با سیستم عصبی محیطی انواع مختلفی دارند.
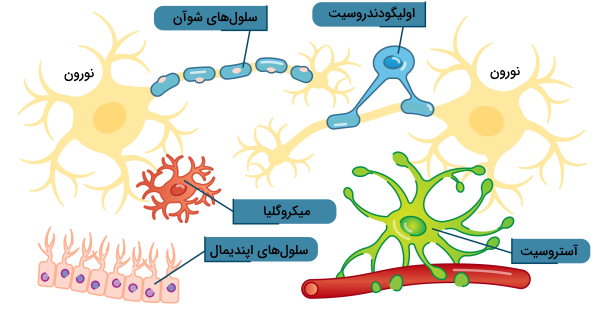
یاخته های پشتیبان سیستم عصبی مرکزی
یاخته های پشتیبان بافت عصبی مرکزی، با تنظیم هومئوستازی بافت عصبی، خارج کردن مواد زائد و تشکیل میلین از نورونها محافظت میکنند. این سلولها از نورونها کوچکتر هستند اما تعداد آنها بسیار بیشتر از نورونها است. در پستانداران مختلف ۳۳ تا ۶۶٪ وزن کلی مغز را یاخته های پشتیبان تشکیل میدهند. پنج نوع سلول پشتیبان در سیستم عصبی مرکزی پستانداران وجود دارد.
- «آستروسیت» (Astrocytes)
- «اولیگودندروسیت» (oligodendrocytes)
- «میکروگلیا» (Microglial)
- «سلولهای اپندیمال» (Ependymal cells)
- «سلولهای گلیال شعاعی» (Radial Glia)
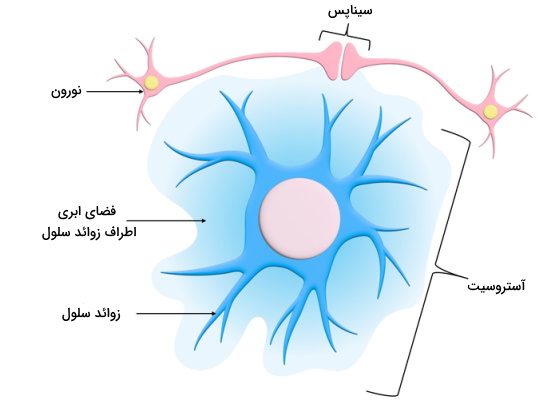
آستروسیتها
آستروسیتها فراوانترین یاختههای پشتیبان در مغز و نیمی از تمام یاخته های عصبی بافت عصبی مرکزی هستند. دلیل نامگذاری این سلولها ساختار ستارهای (-Astro) آنها است. آستروسیتها با تنظیم غلظت انتقالهای دهندههای عصبی در سیناپسها، شرایط محیطی نورونها را تنظیم میکنند. ویژگی شاخص آستروسیتها وجود رشتههای پروتئینی اسیدی (Glial Fibrillary Acidic Proteins | GFAP) در سیتوپلاسم است. این پروتئینها احتمالا در تعیین شکل، حرکت و ارتباط بین آستروسیتها نقش دارند. این سلولها فرایندهای زیر را در مغز و نخاع تنظیم میکنند.
- غلظت انتقالدهنده عصبی را تشخیص میدهند و با ترشح مولکولهای تنظیمی فعالیت نورون را تنظیم میکنند.
- پس از انتقال پیام عصبی، مولکولهای انتقالدهنده عصبی را از سیناپس خارج میکند.
- باقیمانده سلولهای عصبی پس از مرگ را از ماتریکس خارج سلولی خارج میکنند.
- آستروسیتها از اجزای اصلی سد خونی-مغزی هستند که با ترشح مولکولهای تنظیمی، تشکیل اتصالات محکم در در اندوتلیال مویرگها را تحریک و جریان خون به مغز را تنظیم میکنند.
- این یاخته های پشتیبان گلوکز لازم برای فعالیتهای نورونها را ذخیره میکنند.
- آستروسیتها با تنظیم جریان خون محیطی در مغز، اکسیژنرسانی به سلولهای عصبی را تنظیم میکنند.
اولیگودندروسیتها
اولیگوندروسیتها گروهی دیگر از یاخته های پشتیبان در سیستم عصبی مرکزی هستند. ساختار این سلولها از یک کره تشکیل شده است که میلههای غشایی به آن متصل میشوند و در انتهای هر میله غشای سفید و درخشانی قرار دارد. این سلولها دور آکسون میپیچند و غشای محافظتی به نام غلاف میلین ایجاد میکنند. غلاف میلین ساختاری از مولکولهای چربی است که مثل پوشش کابلهای برق، نورونها را از محیط اطراف آنها جدا میکند. این غلاف سرعت انتقال پیام الکتریکی در آکسون را افزایش میدهد.
میکروگلیا
میکروگلیا سلولهای بیضی با زوائد سیتوپلاسمی بسیار و از دسته فاگوسیتها هستند. وظیفه اصلی این یاخته های پشتیبان پاسخ به آسیبها و بیماریهای مغز و نخاع است. این سلولها پس از ایجاد زخم به محل آسیب مهاجرت و با هضم سلول مرده یا خارج کردن پاتوژنها و توکسینها از بافت عصبی محافظت میکنند. به همین دلیل به میرکوگلیا، سلولهای ایمنی بافت عصبی گفته میشود.
سلولهای اپندیمال
سلولهای اپندیمال، یاختههای استوانهای شکلی هستند که از کنار هم قرار گرفتن آنها غشای «اپنیدما» (Ependyma) در مغز و نخاع تشکیل میشود. حرکت منظم مژکهای سلولهای اپندیمال، با حرکت دادن مایع مغزی-نخاعی به توزیع مواد غذایی و دور شدن مواد زائد از سلولها کمک میکند. این سلولها تولید و ترشح مایع مغزی-نخاعی را نیز بر عهده دارند.
سلولهای گلیال شعاعی
این یاختههای پشتیبان در دسته سلولهای بنیادی قرار میگیرند. این سلولها پس از تقسیم و تمایز، سلولهای پشتیبان دیگر ازجمله آستروسیتها و اولیگودندروسیتها را به وجود میآورند. در بسیاری از تحقیقات علوم اعصاب از این سلولها برای بررسی آسیبها و بیماریهای سیستم عصبی استفاده میشود.
یاخته های پشتیبان سیستم عصبی محیطی
نورونهای سیستم عصبی محیطی مانند نورونهاتی مغز و نخاع برای انجام فعالیتهای خود به یاختههای پشتیبان وابسته هستند. هر یک از این یاختهها ساختار، عملکرد و مورفولوژی متفاوتی دارند که برای حمایت از نورونها تخصصیافته مییابد. «سلولهای شوآن» (Schwann cells)، «سلولهای گلیال رودهای» (Enteric Glial Cells) و «گلیالهای ماهوارهای» (Satellite Glial Cells) انواع یاخته های پشتیبان در سیستم عصبی محیطی را تشکیل میدهند.
سلولهای شوآن
این سلولها همتای آستروسیتهای سیستم عصبی مرکزی هستند و ساخت غلاف میلین بر عهده آنها است. غشای پلاسمایی این سلولها دور آکسون یاخته های عصبی میپیچید و با ایجاد یک لایه چربی سرعت انتقال جریان الکتریکی از جسم سلولی به پایان آکسون را افزایش میدهد. سلولهای شوآن غیرمیلینی، دسته دیگری از سلولهای پشتیبان هستند که بدون پوشاندن آکسون از آسیب فیزیکی به این بخش نورون جلوگیری میکنند. برخلاف آستروسیتها، هر سلول شوآن در تشکیل غلاف میلین یک آکسون شرکت میکند. یکی از وظایف سلولهای شوآن خارج کردن باقیمانده سلولهای مرده و ترمیم بخشهای آسیبدیده آکسون است.
سلولهای گلیال رودهای
همانطور که از اسم این سلولها مشخص است، سلولهای گلیای رودهای پشتیبانی نورونهای لوله گوارش را بر عهده دارند. شش نوع مختلف از این نوع یاخته پشتیبان در روده انسان وجود دارد که به عملکرد بهینه نورونهای سمپاتیک و پاراسمپاتیک در این اندام کمک میکنند.
سلولهای ماهوارهای
سلولهای ماهوارهای یاختههای کوچک محافظی هستند که از نورونهای گیرندههای حسی و گانگلیونهای سیستم عصبی سمپاتیک و پاراسمپاتیک پشتیبانی میکنند. وظیفه اصلی این سلولها مثل سایر یاختههای پشتیبان تنظیم هومئوستازی بافت عصبی و جلوگیری از آسیب فیزیکی نورونها با جذب توکسینهای محیط است. سلولهای ماهوارهای همتای میکروگلیای سیستم عصبی مرکزی هستند.
نقش یاخته های پشتیبان در بافت عصبی
یاختههای پشتیبان، سلولهای حمایتی بافت عصبی هستند که چند نقش اصلی دارند.
- تشکیل غلاف میلین اطراف آکسون
- ذخیره گلوکز
- تنظیم خونرسانی و غلظت اکسیژن نورونها
- پاکسازی بافت عصبی از پاتوژنها
بیماریهای یاختههای عصبی پشتیبان
از آنجایی که وظیفه اصلی یاختههای پشتیبان حمایت فیزیکی و متابولیکی از نورونهای عصبی است، اختلال در عملکرد هر یک از آنها تعادل بخشی از بافت عصبی را بهم هم میزند و منجر به بیماریهای ادراکی یا حرکتی مختلفی خواهد شد.
سلولهای گلیا در ایجاد «اختلالهای تحلیل اعصاب» (Neurodegenerative Disorder) نقش اصلی دارند. این اختلالها علائم و شدت متفاوتی دارند اما از بین رفتن نورونها یا بخشی از غلاف میلین در سیستم عصبی مرکزی و تجمع پروتئينها در هسته یا سیتوپلاسم نورون ازجمله علائم مشترک بین آنها است. برای مثال «سندروم گوئیلان بار» (Guillain-Barre Syndrome) یکی از بیماریهای خودایمنی است که سیستم ایمنی بدن غلاف میلین یاخته های بافت عصبی سالم (سلولهای شوآن) را با پاتوژنهای بیماریزا اشتباه میگیرد و آنها را از بین میبرد. این اختلال با بیحسی، از دست حواس پنجگانه در موارد حاد با فلج اندامهای حرکتی همراه است.
یاخته های اصلی بافت عصبی
نورونها یاخته های اصلی بافت عصبی در مهرهداران و بسیاری از بیمهرگان تکاملیافته بعد از رده کیسهتنان هستند. وظیفه اصلی این سلولها برقراری ارتباط بین انواع سلولها و اندامهای مختلف بدن بهوسیله انتقال جریان الکتریکی است. نورونها یاختههای غصبی بسیار تمایزیافتهای هستند که توانایی میتوز و خودترمیمی را از دست دادهاند. تصویر زیر ساختار کلی یک نورون در سیستم عصبی محیطی را نشان میدهد که از سه بخش اصلی تشکیل میشود.
- دندریت: دندریت زوائد سیتوپلاسمی هستند که از جسم سلولی نورون خارج میشوند. ساختار این زوائد معمولا شاخهای است که سبب میشود مساحت سطح بیشتری برای انتقال جریان الکتریکی در اختیار سلول قرار بگیرد. دندریتها پیام را از اندامها و بافتهای مختلف دریافت میکنند و به جسم سلولی انتقال میدهند.
- جسم سلولی: جسم سلولی بین درندریتها و آکسون قرار دارد و بیشتر سیتوپلاسم سلول و اندامکها در این قسمت جای دارند. این سلولها فاقد سانتریول هستند.
- آکسون: آکسون و پایانه آکسون بخشی است که جریان الکترکی را از جسم سلولی دریافت و به نورون یا بافت بعدی منتقل میکند. این بخش از نورون با غلاف میلین پوشیده میشود. بخش سفید سیستم عصبی مرکزی غلاف میلین آکسونها و بخش خاکستری آن جسم سلولی نورون، دندریتها و گرههای رانویه (آکسون بین دو بخش میلیندار) تشکیل شده است.
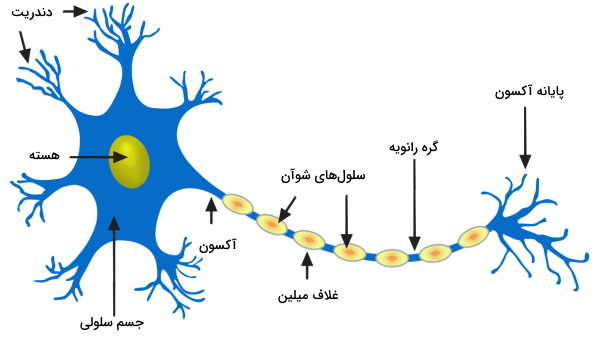
انواع یاخته های اصلی بافت عصبی
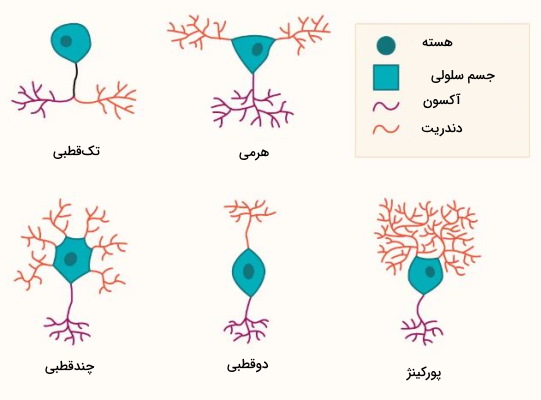
ساختار اصلی همه نورونها از سه بخش دندریت، جسم سلولی و آکسون تشکیل شده است. اما این یاخته های بافت عصبی بر اساس نوع پیامی که منتقل میکنند، شکلهای متفاوتی دارند.
- «نورونهای چندقطبی» (Multipolar neurons): این نورونها از یک آکسون و چند دندریت متقارن تشکیل شدهاند. بیشتر نورونهای سیستم عصبی مرکزی از این نوع هستند.
- «نورونهای تکقطبی» (Unipolar neurons): در این نورونها یک زائده از جسم سلولی خارج میشود که دندریت و آکسون از آن شاخه میگیرند. نورونهای تکقطبی یاختههای عصبی بیمهرگان هستند.
- «نورونهای دوقطبی» (Bipolar neurons): در این نورونهادو زائده سیتودلاسمی در دو جهت مخالف از جسم سلولی خارج میشود. دندریتها در انتهای یکی از این زوايد و آکسون در انتهای دیگر تشکیل خواهد شد. بیشتر نورونهای موجود در شبکیه چشم و بخشی از نورونهای سیستم بویایی و شنوایی از این نوع هستند.
- «نورونهای هرمی» (Pyramidal neurons): این نورونها از یک آکسون و چند شاخه دندریتی هرمی تشکیل شدهاند. بیشتر نورونهای بخش قشری مغز از این نوع هستند.
- «نورونهای پورکینژ» (Purkinje neurons): این یاختههای عصبی یک آکسون و شاخههای دندریتی بسیار زیادی دارند. این نورونها نقش مهاری دارند و از ایجاد پتانسیل عمل در نورون پسسیناپسی جلوگیری میکنند.
یکی دیگر از روشهای تقسیمبندی، بر اساس عملکرد نورونها است. بر این نورونهای نخاع در سه دسته «حسی» (Sensory)، «حرکتی» (Motor) و «بینابینی» (Interneurons) قرار میگیرند.
- نورونهای حسی: این نورونها دندریت بلند و آکسون کوتاهی دارند. نورونهای حسی یا آوران، انواع گیرندههایی هستند که در گوش، پوست، بینی، چشم و زبان تغییرات فیزیکی (دما، فشار و شدت نور) و شیمیایی (مولکولهای بو و مزه) محیط را تشخیص میدهند. این گیرندهها «نورونهای تکقطبی کاذب» (Pseudounipolar) هستند. گیرندههای حسی به چهار نوع اصلی تقسیم میشوند.
- شیمیایی: بیشتر این گیرندهها در جوانه چشایی زبان و سلولهای بویایی بینی قرار دارند.
- دما: این گیرندهها در پوست نسبت به سایر اندامها بیشتر است.
- نور: گیرندههای نوری سلولهای اصلی تشکیلدهنده شبکیه چشم هستند.
- فشار: این گیرندهها تغییرات فشار پوست و خون را تشخیص میدهند.
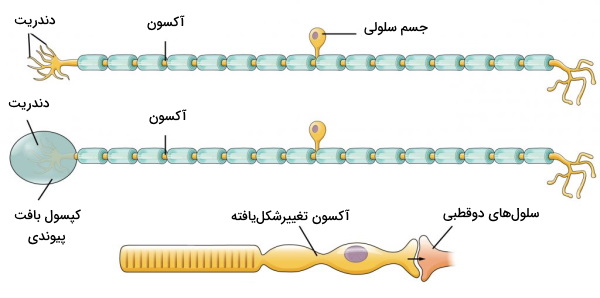
گیرندههای حسی نورونهای تمایزیافته اندامهای حسی هستند. به این نکته توجه کنید که دندریت و آکسون از یک سمت جسم سلولی خارج شدهاند.
- نورونهای حرکتی: این نورونها آکسون بلند و دندریت کوتاهی دارند. نورونهای حرکتی یا وابران، از نخاع شروع میشوند و پیام عصبی را به ماهیچههای صاف و مخطط، غدد و دیگر اندامهای بدن انتقال میدهند. این نورونها از انواع چندقطبی هستند و به دو دسته کلی تقسیم میشوند.
- نورونهای بالایی: این نورنها بین مغز و نخاع ارتباط برقرار میکنند.
- نورونهای پایینی: این نورونها بین نخاع و ماهیچهها ارتباط برقرار میکنند.
- نورونهای بینابینی: دندریت این نورونها کوتاه است اما آکسون ممکن است بلند یا کوتاه باشد. نورونهای بینابینی برقراری ارتباط بین نورونهای حرکتی و حسی بر عهده این دستهاز نورونها است. این یاختههای عصبی مثل نورونهای حرکتی از انواع چندقطبی هستند.
انواع یاخته های اصلی در مغز
تقسیمبندی نورونهای مغز پیچدهتر از نورونهای محیطی و نخاع است. در تقسیمبندی کلی پیامهای عصبی سیستم حسی (قشر شنوایی و بویایی) و پیامهای عصبی سیستم حرکتی (مخچه و قشر حرکتی) به بخشهای جداگانهای از مغز منتقل میشود. اما هر بخش از صدها نورون مختلف تشکیل شده است. انتقالدهندههای عصبی یکی از روشهایی است که عصبشناسان برای تقسیمبندی نورونها از آن بهره میبرند. برای مثال نورونهای GABA گروهی از یاخته های بافت عصبی هستند که بهوسیله انتقالدهنده عصبی آمینوبوتیریک اسید پیام خود را به سلولهای بعدی منتقل میکند.
انتقال پیام در یاخته های اصلی بافت عصبی
در بخشهای قبلی اشاره کردیم که انتقال پیام در نورونها بهوسیله انتقال جریان الکتریکی است. انتقال جریان الکتریکی از یک نورون به سلول دیگر (نورون، ماهیچه و غدد اندوکرین) از راه سیناپس و به کمک انتقالدهندههای عصبی انجام میشود.
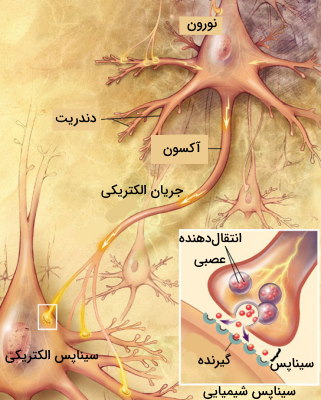
سیناپس
سیناپس مجموعه انتهای پیشسیناپسی، شکاف سیناپس و انتهای پسسیناپسی است. دو نوع سیناپس الکتریکی و شیمیایی انتقال پیام بین یاخته های اصلی بافت عصبی را بر عهده دارند.
- سیناپس شیمیایی: در این سیناپسها آزاد شدن انتقالدهنده عصبی از نورون پیشسیناپسی و اتصال آن به گیرندههای کانالی سلول پسسیناپسی، سبب تغییر پتانسیل عمل و انتقال پیام میشود. انتقالدهندههای عصبی دو اثر مهاری و تحریکی دارند.
- سیناپس الکتریکی: این سیناپس بین دو نورونی ایجاد میشود که بین آنها «اتصالات شکافدار» (Gap Junction) وجود دارد. این سیناپسها کوچکتر از سیناپسهای شیمیایی هستند و فعالیت تحریکی دارند. غشای پلاسمایی نورون در این سیناپسها کانالهای یونی فراوانی دارد که به انتقال سریع پیام کمک میکند. اثر تحریک در این سیناپسها زودتر از سیناپسهای شیمیایی از بین میرود.
انتقال دهنده عصبی
انتقالدهندههای عصبی مولکولهای شیمیایی هستند که مثل هورمونها در انتقال پیام بین سلولهای موجودات زنده نقش دارند. با این تفاوت که هورمونها مولکولهای پپتیدی یا استروئیدی هستند و با جریان خون به اندامهای مختلف منتقل میشوند و عملکرد سلول را تغییر میدهند. اما انتقالدهندههای عصبی مولکولهای کوچکی هستند که در فضای کوچک سیناپس بین دو نورون عصبی یا نورون با سلولهای ماهیچهای و غدد درونریز ارتباط برقرار میکنند. انتقالدهندههای عصبی برخلاف ساختار متفاوت، مکانسیم عمل مشابهی دارند.
- پتانسیل عمل در پایانه آکسون خروج انتقالدهنده عصبی بهوسیله اگزوسیتوز از غشای نورون پیشسیناپسی به شکاف بینسیناپسی را تحریک میکند.
- انتقالدهنده عصبی به گیرنده خود در غشای سلول پسسیناپسی متصل میشوند.
- با بسته (مهار) و باز کردن (تحریک) کانال به طور مستقیم (گیرنده کانالی) و غیر مستقیم (گیرنده غیرکانالی) پتانسیل عمل سلول پسسیناپسی را تغییر میدهد و پیام الکتریکی را منتقل میکند.
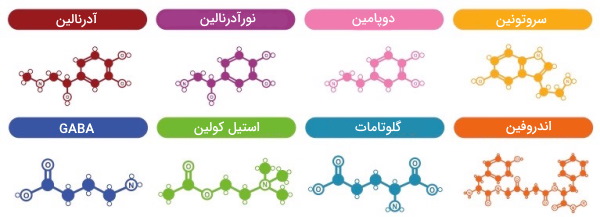
انواع انتقالدهنده عصبی
انتقالدهندههای عصبی از انواع مختلف ساختارهای شیمیایی ازجمله اسید آمینه تغییریافته، پپتید، پورینها، آمین الکلها یا مولکولهای گازی تشکیل شده است.
آمینواسید
آمینواسیدها مولکولهای آلی هستند که علاوه بر شرکت در ساختار پروتئین نقشهای دیگر در بدن زنده ایفا میکنند. تعدادی از این مولکولها به انتقال پیام بین یاختههای بافت عصبی کمک میکنند.
- گاما-آمینوبوتیریکاسید (GABA): این آمینواسید طبیعی یکی از مهارکنندههای اصلی سیستم عصبی است و در انتقال پیامهای بینایی، تنظیم حرکتی و تنظیم اضطراب نقش دارد. این نوروترنسمیتر با اثر آنزیم گلوتامات دکربوکسیلاز بر آمینواسید گلوتامات در پایانه آکسون تولید و ذخیره میشود. GABA انتقالدهنده عصبی اصلی در سیناپسهای مهاری مغز است. داروهای بنزودیازپین که برای کنترل اضطراب تجویز میشود، ترشح GABA را افزایش میدهد.
- گلوتامات: گلوتامات فراوانترین انتقالدهنده عصبی سیستم عصبی است که در فرایندهای شناختی ازجمله حافظه و یادگیری نقش دارد. گلوتامات نوروترنسمیتر سیناپسهای تحریکی است که در پایانه آکسون نورون پیشسیناپسی و با فعالیت آنزیم گلوتامیناز تولید میشود. غلظت زیاد این آمینواسید در سیستم عصب منجر به بیشتحریکی نورون و مرگ سلول میشود. در بیماران مبتلا به آلزایمر، سکته مغزی و تشنج صرع تولید گلوتامات افزایش مییابد.
- گلایسن: گلایسن انتقالدهنده آمینواسیدی و مهاری نخاع است که از تغییر آمینواسید سرین بهوسیله سرین هیدروکسیمتیل ترانسفراز در پایانه عصبی آکسون تولید و ذخیره میشود.
پپتید
مولکولهای زیستی هستند که از کنار هم قرار گرفتن کمتر از ۱۰۰ زیرواحد آمینواسیدی تشکیل میشوند. این مولکولها به انتقال پیام بین یاختههای بافت عصبی کمک میکنند.
- اُکسیتوسین: این پپتید یک نوروهورمون است (هورمونهایی که در نورونها تولید و با جریان خون به سایر اندامها منتقل میشوند) که بهوسیله سلولهای هیپوتالاموس تولید میشود و در تنظیم فرایندهای شناخت اجتماعی، برقراری ارتباط و تولید مثل نقش دارد.
- اندورفین: این پپتیدها انتقالدهندههای عصبی مهاری هستند که با مهار پیام درد، احساس سرخوشی را افزایش میدهند. انجام تمرینات بیهوازی تولید اندورفین در یاختههای بافت عصبی را افزایش میدهد.
نوروترنسمیترهای پپتیدی تنها گروه از مولکولهای پیامرسان سیستم عصبی هستند در جسم سلولی تولید و بهوسیله وزیکولهای حامل به پایانه آکسون منتقل میشوند. پس از رونویسی و ترجمه، این پپتیدها وارد دستگاه گلژی جسم یاختهای و پس از آن به وسیله میکروتوبولهای آکسون به پایانه منتقل میشوند. به همین دلیل انتقال پیامهای وابسته به نروترنسمیترهای پپتیدی آهستهتر از انتقالدهندههای دیگر است.

کتکولآمین یا مونوآمینها
این گروه از انتقالدهندههای عصبی مولکولهای آلی هستند که از اتصال گروه آمین به حلقه آروماتیک و دو اتم کربن تشکیل میشوند. نورونها این ترکیبات را از آمینواسیدهای آروماتیک ازجمله فنیلآلانین، تیروزین و تریپتوفان و بهوسیله آنزیم دکربوکسیلاز تولید میکنند.
- اپینفرین: اپینفرین یا آدرنالین مولکول پیامرسانی است که هورمون غدد فوق کلیه و انتقالدهنده عصبی یاختههای عصبی مغز است.
- نوراپینفرین: پس از تشکیل وزیکولهای سیناپسی حاوی دوپامین، آنزیم غشایی دوپامینبتا هیدروکسیلاز، دوپامین را به نوراپینفرین تبدیل میکند. در نتیجه برخلاف سایر نروترنسمیترهای غیر پپتیدی، نورآدرنالین بهوسیله واکنشهای آنزیمی سیتوپلاسم تولید نمیشود. این انتقالدهنده عصبی در پاسخهای حمله یا فرار نقش دارد. این انتقالدهنده عصبی اندامهای حرکتی را برای فرار و نورونهای مغز را برای تحلیل شرایط و پاسخ به موقع به خطر یا استرس تحریک میکند. غلظت نوراپینفرین بافت عصبی در زمان خواب به کمترین میزان و در مواقع استرس به بیشترین میزان میرسد.
- هیستامین: هیستامین انتقالدهنده عصبی نورونهای مغز و نخاع است که پس از تغییر آمینواسید هیستیدین بهوسیله آنزیم دکربوکسیلاز در پایانه آکسون تولید و ذخیره میشود. به علاوه ماستسلها در پاسخ به آلرژنها هیستامین ترشح میکنند.
- دوپامین: دوپامین یکی از انتقالدهندههای عصبی است که در پایانه آکسون نورون پیشسیناپسی و در یک فرایند دو رمحلهای از تغییر آمینواسید تیروزین تولید و ذخیره میشود. این نوروترنسمیتر در انتقال پیامهای پاداش و انگیزه نقش دارد. بسیاری از مخدرها با غلظت دوپامین در بافت عصبی را افزایش میدهند. به علاوه، دوپامین در هماهنگی اندامهای حرکتی بدن نقش دارد. پارکینسون یکی از بیماری تحلیل یاخته های عصبی است که منجر به اختلال در عملکرد حرکتی بیمار میشود. دلیل ایجاد این بیماری از بین رفتن نورونهای ترشحکننده دوپامین در مغز است.
- سروتونین: سروتونین یکی از مهمترین هورمونها و انتقالدهندههای عصبی بدن برای تنظیم خلق، خواب، اضطراب، میل جنسی و اشتها است. این نوروترنسمیتر در یک مسیر دو مرحلهای از آمینواسید تریپتوفان در پایانه آکسون تولید میشود. در این مسیر تریپتوفان بهوسیله آنزیم هیدروکسیلاز و سپس دکربوکسیلاز ال-آمینواسید آروماتیک به سروتونین تبدیل میشود.
مهارکنندههای انتخابی بازجذب سروتونین (SSRIs) دستهای ترکیبات شیمیایی هستند که با مهار بازجذب سروتونین در مغز سبب تغییر خلق و کاهش اضطراب میشوند. - اپینفرین: اپینفرین یکی از مولکولهای انتقال پیام مشترک بین سیستم تنظیم هورمونی و عصبی است. تعداد بسیار کمی از نورونها از این نوروترنسمیتر برای انتقال پیام عصبی بهره میبرند. برای سنتز این انتقالدهنده عصبی در پایانه آکسون باید نوراپینفرین تولید شده در ویزیکولهای ذخیرهای وارد سیتوپلاسم شود. سپس آنزیم فنیل اتانول آمین-N-متیل ترانسفراز، نوراپینفرین را به اپی نفرین تبدیل میکند.
پورینها
پورینها بازهای آلی هستند که نقش اصلی آنها تشکیل نوکلئوتید و شرکت در ساختار اسیدنوکلئيکها است. اما در انتقال برخی پیامهای عصبی نیز نقش دارند.
- آدنوزین: آدونوزین مولکولی است که از پیوند باز آدنین با قند ریبوز تشکیل میشود. این مولکول یکی از انتقالدهندههای عصبی یاختههای عصبی مغز است که در افزایش کیفیت خواب نقش دارد.
- آدنوزین تریفسفات (ATP): ATP مولکول ذخیرهکننده انرژی سلولها و انتقالدهنده عصبی سیستم عصبی مرکزی و محیطی است. این انتقالدهنده عصبی به انتقال پیامهای ارادی، حسی و ارتباط نورونها با یاختههای پشتیبان کمک میکند.
مولکولهای گاز
برخی نورونها برای انتقال پیامهای خود به کمک مولکولهای گازی کوچک نیاز دارند.
- نیتریک اکسید (NO): این انتقالدهنده عصبی با کاهش پتانسیل الکتریکی ماهیچههای صاف سبب انبساط ماهیچه، افزایش قطر رگ و جریان خون به بافتها میشود.
- کربن مونوکسید (CO): کربن مونوکسید گازی بیرنگ، بیبو و بسیار خطرناک برای انسان است. اما در به طور طبیعی در برخی یاختههای عصبی بدن تولید میشود و پاسخهای التهابی را تنظیم میکند.
استیلکولین
استیلکولین از ایجاد پیوند استری بین استیکاسید و کولین (الکل آمینی) در پایانه آکسون و بهوسیله آنزیم کولین استیل ترانسفراز تشکیل و در وزیکولهای انتقالدهنده ذخیره میشود. استیل کولین نروترنسمیتر اصلی در سیستم عصبی پاراسمپاتیک و سیناپس ماهیچه است. استیلکولین میتواند سلول پسسیناپسی را مهار یا تحریک کند. در نورونهای حرکتی ماهیچه، استیلکولین در وزیکولهای پایانه آکسون ذخیره میشود. انتقال پتانسیل عمل به پایانه آکسون، سبب آزاد شدن استیل کولین در شکاف سیناپسی و اتصال آن به گیرندههای کانالی سلول پسسیناپسی خواهد شد. باز شدن کانالها با ورود یون سدیم به سلول ماهیچهای و افزایش پتانسیل عمل این سلولها، انقباض ماهیچه را به همراه خواهد داشت. این انتقالدهنده در سیستم عصبی مرکزی و تنظیم فرایندهای یادگیری نیز نقش دارد.

انتقال دهنده عصبی چگونه وارد فضای سیناپسی می شود ؟
وزیکولهای انتقالدهنده عصبی در پایانه آکسون نورونهای پیشسیناپسی وجود دارد. زمانی که پتانسیل عمل به انتهای آکسون میرسد، کانالهای ولتاژی پایانه آکسون باز و یون کلسیم وارد پایانه آکسون میشود. یون کلسیم به فرایند اگزوسیتوز انتقالدهندههای عصبی کمک میکند. برهمکنش انتقالدهنده عصبی با گیرنده اختصاصی خود در غشای سلول پسسیناپسی تغییر پتانسیل عمل و انتقال جریان الکتریکی به سلول پسسیناپسی را به دنبال دارد.
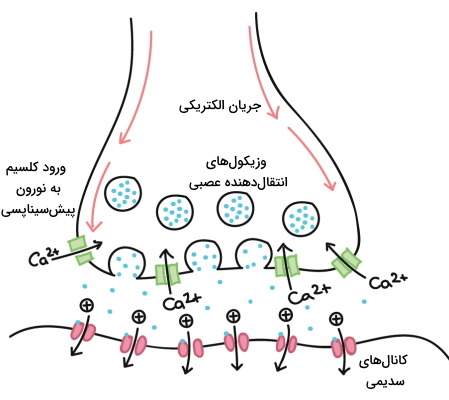
گیرنده انتقال دهنده عصبی
گیرنده انتقالدهندههای عصبی پروتئينهای عرض غشایی هستند که به دو گروه تقسیم میشوند.
- کانالهای یونی: اتصال انتقالدهنده عصبی به این گیرندهها سبب باز شدن کانال و ورود یونها به سیتوپلاسم میشود. گیرنده استیلکولین در غشای ماهیچه اسکلتی از این گروه است.
- گیرندههای غیر یونی: اتصال انتقالدهنده عصبی به این گیرندهها با فعال کردن پیامبر ثانویه، کانالهای دریچهدار را باز و در غشا پتانسیل عمل ایجاد میکند. پیام عصبی بهوسیله این گیرندهها آهستهتر منتقل میشود. گیرنده استیلکولین در غشای ماهیچه قلب از این گروه است.
غیر فعال شدن انتقال دهنده عصبی
انتقالدهندههای عصبی پس از فعال یا مهار کردن سلول پسسیناپسی به شکاف سیناپسی برمیگردد. برای جلوگیری از بیشتحریکی سلول پسسیناپسی، این مولکول باید سریع از سیناپس خارج شود. انتقالدهندههای عصبی با سه مکانیسم زیر از سیناپس خارج میشوند.
- تجزیه: ساختار انتقالدهنده عصبی بهوسیله آنزیم تغییر میکند و نمیتواند به گیرنده متصل شود. استیلکولین استراز آنزیمی است که استیلکولین را در شکاف سیناپسی هیدرولیز میکند. استات حاصل از این واکنش آنزیمی وارد سلولهای اطراف میشود و کولین به نورون پیشسیناپسی برمیگردد.
- انتشار: انتقالدهنده عصبی بهوسیله انتشار وارد سلولهای پشتیبان و از دسترس گیرنده خارج میشود.
- بازجذب: ساختار مولکول انتقالدهنده بدون تغییر باقی میماند و بهوسیله ناقلهای پروتئينی موجود در غشای نورون پیشسیناپسی بازجذب میشود. حذف سروتونین از سیناپس از این راه انجام میشود.
اثر دارو بر انتقال دهنده عصبی
بسیاری از داروهای درمان بیماریها و اختلالهای عصبی، همچنین بسیاری از مخدرها با فعالیت نورونها را با تغییر عملکرد انتقالدهندههای عصبی، تغییر میدهند. این داروها ساختار و اثر متفاوتی دارند.
- ساختار آگونیست و آنتاگونیست: داروهای آگونیست اثر انتقالدهندههای عصبی را افزایش میدهند و آنتاگونیستها انتقالدهندههای عصبی را مهار میکنند. بسیاری از مخدرها اثر آگونیست دارند.
- اثر مستقیم و غیرمستقیم: بعضی از داروهای ساختار مشابه انتقالدهندههای عصبی دارند و به جای این مولکولها در انتقال پیام شرکت میکنند. دسته دیگر داروها با اثر بر گیرندههای انتقالدهنده عصبی، به طور غیرمستقیم فعالیت نورونها را تغییر میدهند. برای مثال «پاکسیل» (Paxil) با مهار ناقلهای پروتئین در غشای نورون پیشسیناپسی، غلظت سروتونین در مغز را افزایش میدهد.
پتانسیل عمل یاخته های بافت عصبی
انتقال پیام بین نورونها و نورونها با سلولهای دیگر وابسته به ایجاد جریان الکتریکی در این سلولها است. به همین دلیل غشای پایانه آکسون هر نورون محل تجمع تعدادی زیادی کانال یونی است که به انتقال جریان کمک میکنند. سیتوپلاسم سلول به دلیل غلظت بیشتر یونهای منفی، معمولا بار منفی بیشتری نسبت به ماتریکس خارج سلولی دارد. به همین دلیل پتانسیل غشا در حالت استراحت است. پتانسیل الکتریکی غشای آکسون ثابت نیست و در اثر انتقال پیام الکتریکی مثبتتر یا منفیتر میشود. انتقال پیام بین نورونها بهوسیله تغییر ناگهانی پتانسیل الکتریکی غشا پس از باز شدن کانالهای دریچهدار سدیمی انجام میشود. این فرایند ایجاد پتانسیل عمل نام دارد. نمودار زیر مراحل ایجاد پتانسیل عمل را نشان میدهد.
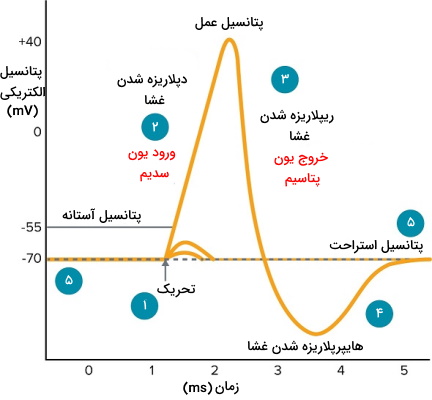
- اتصال انتقالدهنده عصبی به رسپتور سبب باز شدن کانال دریچهدار سدیمی، ورود یون سدیم مثبت به سیتوپلاسم نورون و افزایش پتانسیل الکتریکی غشا میشود.
- تداوم اثر محرک و جریان بونهای سدیم مثبت به سیتوپلاسم سلول سبب میشود پتانسیل الکتریکی غشا به بسیار مثبت () و غشا دپلاریزه شود.
- برای حفظ هومئوستازی سلول، کانالهای درچهدار سدیمی غیرفعال و کانالهای دریچهدار پتاسیمی باز میشوند. در نتیجه ورود سدیم به سلول مهار و با خروج یون پتاسیم، پتانسیل الکتریکی غشا منفی خواهد شد.
- با خروج یون پتاسیم و توقف ورود یون سدیم پتانسیل الکتریکی غشا به کمتر از حالت استراحت کاهش پیدا میکند و غشا هایپرپلاریزه میشود.
- با بسته شدن کانالهای دریچهدار پتاسیمی و فعالیت پمپ سدیم-پتاسیم، پتانسیل غشا به حالت استراحت برمیگردد.
نقش کانال های یونی غشای یاخته های عصبی در ایجاد پتانسیل عمل چیست ؟
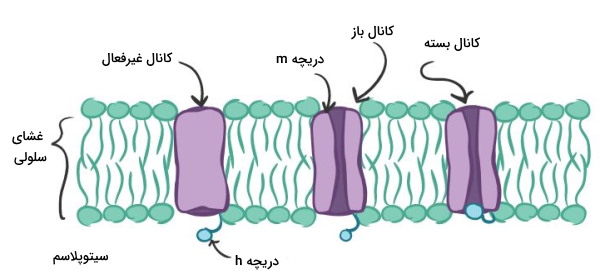
پتانسیل عمل چیزی جز تغییر متناوب پتانسیل الکتریکی غشای یاختههای عصبی به دلیل باز و بسته شدن کانالهای غشایی و تغییر جریان ورودی و خروجی یونها نیست. وضعیت این کانالها در حالت استراحت و پتانسیل عمل با هم متفاوت است.
- پتانسیل استراحت: در این حالت همه کانالهای دریچهدار سدیمی و پتاسیمی بسته هستند. کانالهای نشتی سدیم و پتاسیم پتانسیل استراحت یاخته های عصبی را تنظیم میکنند.
- پتانسیل عمل: ایجاد پتانسیل عمل وابسته به فعالیت کانالهای دریچهدار سدیمی و پتاسیمی است.
- کانالهای سدیمی: این کانالها دو دریچه دارند. دریچه m زمانی باز میشود که پتانسیل غشا به حد آستانه برسد. دریچه h معمولا باز است و با دپلاریزه شدن غشا بسته میشود. در حالت استراحت دریچه m بسته است و یون سدیم وارد سیتوپلاسم نمیشود. با تغییر پتانسیل الکتریکی غشا دریچه m باز و یون سدیم وارد سیتوپلاسم میشود. با دپلاریزه شدن غشا دریچه h بسته و ورود یون سدیم به سیتوپلاسم متوقف میشود.
- کانالهای پتاسیمی: دریچه n معمولا بسته است و با دپلاریزه شدن غشا به آرامی باز میشود.
جمع بندی
بافت عصبی مجموعهای از نورونهای انتقالدهنده پیام و سلولهای پشتیبانی است که از آنها محافظت میکنند. این سلولها در دو بخش اصلی سیستم عصبی مرکزی شامل مغز و نخاع و سیستم عصبی محیطی شامل اندامهای حسی، عصب سمپاتیک و پاراسمپاتیک قرار دارند. در این مطلب با انواع یاختههای پشتیبان و عملکرد متفاوت آنها با نورونها آشنا شدیم. به علاوه یاخته های اصلی بافت عصبی که نورونها هستند و چگونگی انتقال جریان الکتریکی در آنها را بررسی کردیم.










سلام عالی بود ممنون از شما
ولی اینجا رو تصحیح کنید دریچه m وقتی به حد آستانه میرسه باز میشه نه وقتی مثبت میشه
حد آستانه حدود منفی ۶۵ میلی ولت هست
با سلام و وقت بخیر،
مطلب بازبینی و اصلاح شد.
از دقت و همراهی شما با مجله فرادرس سپاسگزاریم.