



گوارش در روده – به زبان ساده
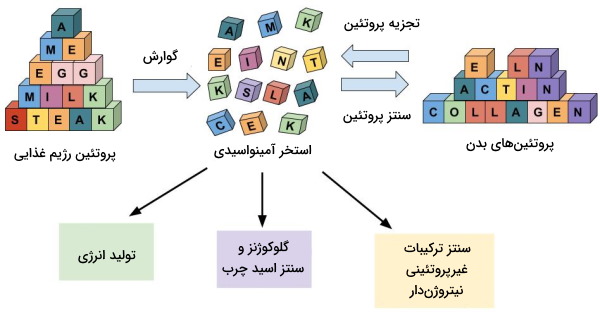
گوارش فرایند تبدیل پلیمرهای بزرگ و تودهای غذا به زیرواحدهای کوجک سازنده آنها است. این فرایند به دو صورت مکانیکی و شیمیایی انجام میشود. گوارش مکانیکی در دهان شروع میشود و دندانها توده غذا را خرد و تکهتکه میکنند. در بخشهای دیگر لوله گوارش حرکات دیواره وظیفه گوارش مکانیکی را بر عهده دارد. گوارش شیمیایی بهوسیله آنزیمهای هیدرولازی، پیوند بین زیرواحدهای پلیمری را تجزیه و مولکولهای قابل جذب تولید میکند. در این مطلب گوارش در روده انسان را به زبان ساده توضیح میدهیم.
- ساختار بخشهای مختلف روده و ویژگیهای هر کدام را میآموزید.
- یاد میگیرید چه تفاوتهایی میان عملکرد روده کوچک و بزرگ در گوارش وجود دارد.
- خواهید آموخت که هورمونها و آنزیمها چه نقشی در تنظیم گوارش دارند.
- نحوه جذب آب، الکترولیت و مواد مغذی در روده را یاد خواهید گرفت.
- با عملکرد سلولهای تخصصی دیواره روده و اهمیت آنها آشنا میشوید.
- میآموزید فرایندهای مکانیکی و شیمیایی گوارش چگونه در روده انجام میشوند.




گوارش در روده
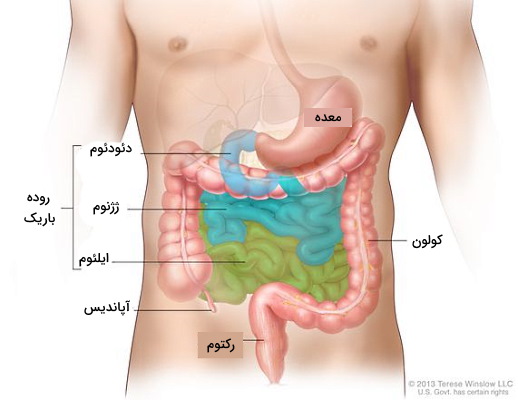
روده کوچک و بزرگ بزرگترین بخشهای لوله گوارش هستند که کل بخش پایینی حفره شکمی را پر میکنند. کیموس معده مخلوطی از غذای نیمهگوارش شده، آنزیم و اسید معده است که با عبور از اسفنکتر پیلوری در انتهای معده، وارد روده باریک میشود. بخش اصلی گوارش کربوهیدراتها و چربیها در این لوله ۳ متری با قطر ۱ سانتیمتر انجام میشود. پس از روده باریک، کیموس وارد روده بزرگ خواهد شد.
این بخش از لوله گوارش بیشتر در جذب آب و یونها و تشکیل مدفوع نقش دارد و فعالیت گوارشی آن بهوسیله باکتریهای مفید (فلور نرمال روده) انجام میشود. برای توضیح گوارش در روده ابتدا آناتومی و بافت این دو اندام را بررسی میکنیم.
ساختار روده باریک
روده باریک به سه بخش تقسیم میشود که از سمت معده (پروکسیمال) تا روده بزرگ (دیستال) «دئودئوم» (Duodenum)، «ژژنوم» (Jejunum) و «ایلئوم» (Ileum) نام دارند.
- دئودئوم: کوتاهترین (۲۵٫۴ سانتیمتر) ناحیه روده کوچک است و پس از اسفنکتر پیلوری روده را شروع و قبل از اتصال به ژژنوم، پیچ S شکلی نزدیک بخش بالایی پانکراس ایجاد میکند. دئودئوم به چهار بخش بالایی، پایینرونده، افقی و بالارونده تقسیم میشود. صفرا و آنزیمهای پانکراس یا لوزالمعده از راه مجرای کبدی و با کنترل اسفنکتر اُدی وارد این بخش از روده باریک میشوند.
- ژژنوم: ژژنوم بخش میانی و بلند (۰٫۹ متری) روده است که دئودئوم را به ایلئوم متصل میکند. اما مرز مشخصی بین این بخش و ایلئوم وجود ندارد.
- ایلئوم: ایلئوم بخش پایانی و بلندترین (۱٫۸ متر) بخش روده باریک است. تعداد رگهای خونی و چینخوردگیهای مخاط دیواره در ایلئوم بیشتر از دئودئوم و ژژنوم است. ایلئوم بهوسیله اسفنکتر ایلئوسیکال با سکوم روده بزرگ ارتباط دارد.
بافت روده باریک
دیواره روده کوچک از چهار لایهای (مخاط، زیرمخاط، ماهیچههای صاف و سروز یا صفاق) تشکیل میشود که در سایر بخشهای لوله گوارش وجود دارند. اما مخاط و زیرمخاط موجود در این بخش سلولهایی دارد که برای گوارش و جذب مواد غذایی تخصصی شدهاند. تاخوردگی حلقوی، ویلی و میکروویلی ویژگیهای ساختاری هستند که مساحت سطح روده را برای جذب مواد افزایش میدهند (بیش از ۶۰۰ برابر) و تعداد آنها در دو-سوم ابتدایی روده بیشتر است.
- تاخوردگی حلقوی: «تاخوردگی» (Fold) یا «چینخوردگی» (Plica) حلقوی، شیارهای عمیق در مخاط و زیرمخاط دیواره روده هستند که از ابتدای دئودئوم شروع میشوند و در میانه ایلئوم پایان مییابد. شکل این ساختارها سبب حرکت مارپیچ کیموس در روده و کاهش سرعت آن میشود. در نتیجه فرصت کافی برای جذب مواد از سلولها فراهم خواهد شد.
- ویلی: ویلیها، ویلوس (مفرد) یا پرز روده، ساختارهای انگشتی سطح چینخوردگیها هستند که رگهای خونی در آنها قرار دارد. در هر میلیمتر مربع اپیتلیوم روده، ۲۰ تا ۴۰ ویلی وجود دارد که مساحت سطح این لایه را افزایش میدهد. هر ویلی از سلولهای جذبکننده، یک سرخرگچه، یک سیاهرگچه و یک رگ لنفاوی (لاکتئال) تشکیل شده است. آمینواسیدها و مونوساکاریدها پس از عبور از سلولهای جذب، مستقیم وارد سرخرگچه میشوند اما اسیدهای چرب و سایر لیپیدها وارد لاکتئال خواهند شد و بهوسیله سیستم لنفاوی وارد خون میشوند.
- میکروویلی: میکروویلیها یا میکروویلوس زوائد سیتوپلاسمی بسیار کوچک (۱ میکرومتر) در سطح سلولهای جذبکننده ویلی هستند که ساختار آنها بهوسیله فیلامنمتهای اسکلت سلولی تنظیم میشود. از آنجایی که تصویر میکروسکوپی این بخش از بافت روده شبیه به برس مسواک است به آن «حاشیه مسواکی» (Brush Border) نیز میگویند. آنزیمهای هیدرولاز کربوهیدراتها و پروتئینها در غشای این بخش از روده باریک قرار دارد. در هر میلیمتر مربع اپیتلیال روده، تقریبا ۲۰۰ میلیون میکروویلی وجود دارد که مساحت سطح این بافت را به مقدار قابل توجهی افزایش میدهد.
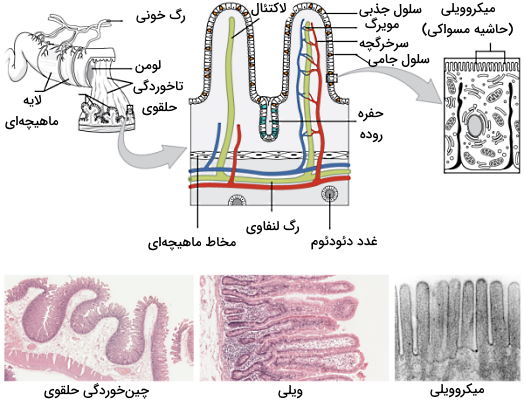
سلول های دیواره روده باریک
علاوه بر ویژگیهای ساختاری که در بخش قبل توضیح دادیم، اپیتلیال روده بین هر دو میکروویلی شکافی دارد که سلولهایی دارد که هورمونها و شیره روده را ترشح میکنند. این بخش از اپیتلیال روده حفره یا غده لیبرکان (Crypts of Lieberkuhn) نام دارد. شیره روده ترکیبی قلیایی (pH ۷٫۴ تا ۷٫۸) از آب و مخاط است که هر روز حدود ۰٫۹۵ تا ۱٫۹ لیتر از آن در پاسخ به حرکات روده و تحلیه کیموس به دئودئوم ترشح میشود. مخاط روده تنها از سلولهای زیرمخاط یا غدد برونر (Brunner’s Glands) ترشح میشوند. گوارش در روده بهوسیله سلولهای مختلفی انجام و تنظیم میشود.
- سلولهای جذبکننده: سلولهای اپیتلیالی هستند که گوارش و جذب مواد کیموس را بر عهده دارند.
- سلولهای جامی: سلولهای اپیتلیال که مخاط روده را ترشح میکنند.
- سلولهای پنز: سلولهای «پنز» (Paneth)، غدد رودهای هستند که آنزیم لیزوزیم ترشح میکنند و فاگوسیتوز باکتریها را انجام میدهند.
- سلولهای G: سلولهای غدد دئودئوم هستند که وظیفه ترشح هورمون گاسترین را بر عهده دارند.
- سلولهای I: سلولهای غدد دئودئوم که با ترشح هورمون کولهسیستوکینین ترشح آنزیمهای پانکراس و صفرا را تحریک میکنند.
- سلولهای K: این سلولها با ترشح پپتید شبه انسولینی تنظیم کننده گلوکز، ترشح انسولین را تحریک میکنند.
- سلولهای M: این سلولها در دئودئوم و ژژنوم قرار دارند و با ترشح هورمون موتیلین، حرکات دودی روده، تخلیه معده و ترشح پپسین از سلولهای اصلی معده را تحریک میکنند.
- سلولهای S: سلولهای ترشحکننده هورمون سکرتین که ترشح اسید معده و pH دئودئوم را تنظیم میکند.
گوارش مکانیکی غذا در روده باریک
دو نوع انقباض ماهیچههای صاف دیواره روده سبب ایجاد حرکات دودی و قطعهقطعه کننده میشوند که به گوارش و جذب غذا کمک میکنند. اگرچه حرکات دودی که در معده به مخلوط شدن شیره معده و غذا کمک میکند در این بخش از لوله گوارش وجود ندارد. انقباض قطعهقطعه کننده ماهیچههای حلقوی یکی از حرکات لوله گوارش است که به گوارش غذا در روده کوچک و بزرگ کمک میکند.
این انقباضها به جای حرکت دادن کیموس در روده، کیموس را با آنزیمهای گوارشی مخلوط میکنند و امکان جذب مونومرها از سلولهای جذبی را افزایش میدهند. بیشترین تعداد انقباض قطعهقطعه کننده در دئودئوم (حدود ۱۲ بار در دقیقه) و کمترین تعداد این انقباض در ایلئوم (حدود ۸ بار در دقیقه) وجود دارد.
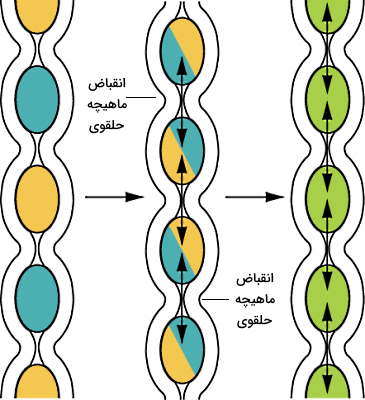
وقتی بیشتر کیموس جذب شد، روده از حالت منبسط خارج میشود و حرکات دودی، جای حرکات قطعهقطعهکننده را میگیرند. ترشح موتیلین از سلولهای M مخاط روده حرکات دودی در روده را تحریک میکند. این حرکات سبب میشوند کیموس کمی در دئودئوم به جلو حرکت کند و سپس متوقف خواهد شد. این حرکات در بازه زمانی منظم تکرار خواهد شد و کیموس را از دئودئوم به انتهای ایلئوم منتقل خواهد کرد. در انتهای ایلئوم حرکات دودی سبب باز شدن اسفنکتر ایلئوم و انتقال کیموس به سکوم روده بزرگ میشود.
گوارش شیمیایی در روده کوچک
گوارش شیمیایی پلیمرهای غذا در دهان و معده شروع میشود و آنزیمهای حاشیه مسواکی روده و پانکراس این فرایند را کامل میکنند. آمیلاز بزاق گوارش کربوهیدرات را در دهان شروع میکند و پپسین معده ساختار پیچیده پروتئینها را به پلیپپتیدهای کوچکتر میشکند. بیشتر گوارش لیپیدها در این بخش از لوله گوارش و به کمک صفرا و لیپاز پانکراس انجام میشود. آنزیمهای گوارش در روده به دو دسته پروتئينهای عرض غشایی (دیساکاریدازها و پپتیداز) و لومنی (انتروکینازها) تقسیم میشوند.
- دیساکاریدازها: این آنزیمها پروتئینهای عرض غشایی میکروویلی هستند که دیساکاریدها اولیگوساکاریدهای منشعب را هیدرولیز میکنند. مالتاز (تجزیه مالتوز به گلوکز)، لاکتاز (هیدرولیز لاکتاز به گالاکتوز و گلوکز) و ساکاراز (هیدرولیز ساکارز به فروکتوز و گلوکز) کربوهیدرازهای حاشیه مسواکی روده هستند.
- پپتیداز: دیپپتیداز، پروتئازی است که در غشای میکروویلیها قرار دارد و پیوند پپتیدی در دیپپتیدها را هیدرولیز میکند. این آمینوپپتیدازها آخرین آنزیمهایی هستند که پروتئينها را برای جذب در روده آماده میکنند.
- انتروکیناز یا انتروپپتیداز: این آنزیم بهوسیله سلولهای غده لیبرکان ترشح میشود. این آنزیم زیموژنهای (آنزیمهای غیر فعال) شیره پانکراس را به آنزیمهای فعال تبدیل میکند.
آنزیمهای پانکراس دسته دوم آنزیمهایی هستند که گوارش مواد در روده را انجام میدهند. این آنزیمها به همراه اجزای دیگر شیره پانکراس (آب و بیکربنات) پس از عبور از مجرای پانکراس به دئودئوم میرسند. بیکربنات موجودر این شیره، اثر اسیدی کیموس را خنثی و محیط را برای فعالیت آنزیمهای لوزالمعده آماده میکند. سه دسته آنزیم در شیره پانکراس وجود دارد.
- آمیلاز: این آنزیم مشابه آنزیم آمیلاز غدد بزاقی است که گوارش شیمیایی کربوهیدرات را در دهان شروع میکند و پیوندهای آلفا گلیکوزیدی در زنجیره کربوهیدراتی اصلی در نشاسته و گلیکوژن را هیدرولیز میکند.
- لیپاز: لیپاز پانکراس دیآسیل گلیسرولهایی که بهوسیله لیپاز معده ایجاد شده است به گلیسرول و اسید چرب هیدرولیز میکند.
- پپتیدازها: پروتئازهای پانکراس نقش اصلی در تبدیل زنجیرههای پلیپپتیدی کوچک به آمینواسیدها و دیپپتیدها دارند. تریپسینوژن، کموتریپسینوژن و کربوکسیپپتیدازها، پروتئازهای شیره لوزالمعده هستند.
- تریپپسینوژن ازجمله زیموژنهایی است که با فعالیت انتروکیناز فعال میشود و پیوندهای پپتیدی در انتهای کربوکسیلی لیزین و آرژینین را میشکند.
- کموتریپسینوژن یکی دیگر از زیموژنهای پانکراس است که بهوسیله تریپسین فعال میشود. کموتریپسین فعال، پیوند پپتیدی بین دو آمینواسیدی را هیدرولیز میکند که یکی از آنها آروماتیک (تریپتوفان، فنیل آلانین و تیروزین) است.
- کربوکسیپپتیدازها، اگزوپپتیدازهایی هستند که اتم روی موجود در جایگاه فعال این پروتئازها به هیدرولیز پیوند پپتیدی در انتهای C زنجیره پلیپپتید کمک میکند.
- الاستاز مانند سایر پروتئازهای دستگاه گوارش به شکل زیموژن از سلولهای آسینار پانکراس ترشح و با واکنش آنزیمی تریپسن فعال میشود. سرین موجود در جایگاه فعال این اندوپپتیداز به هیدرولیز پیوند پپتیدی در الاستین و انواع دیگه پروتئین کمک میکند.
تمام پروتئینهای رژیم غذایی در سه ساختار تریپپتید، دیپپتید و آمینواسید بهوسیله سلولهای دیواره دئودئوم و ابتدای ژژنوم جذب میشوند. این مواد بر اساس نوع زنجیره جانبی در آمینواسید، بهوسیله انتقال فعال و انتشار تسهیل شده از حاشیه مسواکی عبور میکنند و همراه جریان خون (سیاهرگ انتقالی کبد) به کبد منتقل میشوند. ۵۰ تا ۶۰ درصد آمینواسید در کبد باقی میماند و برای سنتز پروتئینهای جدید، بازهای آلی (پورین و پیریمیدین) و تولید انرژی مصرف میشوند. به این نکته توجه کنید که پروتئینها مثل کربوهیدراتها (گلیکوژن کبد و ماهیچهها) و مولکولهای لیپیدی (تریگلیسیرید بافت چربی) در بدن ذخیره نمیشوند و اگر در فرایندهای سنتزی شرکت نکند، به اوره و دیاکسید کربن تبدیل شده و از بدن خارج میشود.
در پایان گوارش کربوهیدراتها در روده باریک، سه مونوساکارید گلوکز، گالاکتوز و فروکتوز تولید میشود. گلوکز و گالاکتوز با انتقال فعال ثانویه و بهوسیله پروتئین همانتقال گلوکز-سدیم (SGLT1) از سطح لومنی، وارد سلولهای میکروویلی میشوند. و با انتشهار تسهیل شده و بهوسیله ناقل پروتئینی GLUT2 وارد خون شده و به کبد منتقل میشوند. ورود و خروج فروکتوز به و از سلولهای دیواره میکروویلی با انتشار تسهیل شده و بهوسیله ناقلهای پروتئینی GLUT5 و GLUT2 انجام میشود.
گوارش چربی در روده باریک
گوارش مکانیکی و شیمیایی چربیها در دهان شروع میشود. این ترکیبات بهوسیله فعالیت دندانها و زبان خرد و تکهتکه میشوند. بهعلاوه بزاق ترکیب امولسیونسازی است که با تشکیل میسلهای لیپیدی و تریگلیسریدی، اثر آنزیم لیپاز زبان در گوارش چربیها را افزایش میدهد. فرایندهای این بخش مولکولهای بزرگ چربی را تبدیل به ذرات کوچکتری میکند که در بخشهای بعدی بهتر تجزیه میشوند.
ذرات چربی پس از عبور از مری و اسفنکتر کاردیا وارد معده میشوند. فعالیت لیپاز معده، تریگلیسرید را به دیآسیل گلیسرول و یک اسید چرب تبدیل میکند. دیآسیل گلیسرول خود به عنوان یک عامل امولسیونساز، تشکیل میسیلهای لیپیدی را افزایش میدهد. ۲ تا ۴ ساعت پس از هر وعده غذایی تنها ۳۰ درصد تریگلیسریدها به دیآسیل تبدیل میشوند و بیشتر چربی بهوسیله لیپاز پانکراس و در روده کوچک هیدرولیز خواهد شد.
ورود کیموس معده با ترکیب چربی زیاد با تحریک هورمون ... ترشح صفرا از کبد را تحریک میکند. صفرا ترکیبی از نمکهای صفرایی (نمکهای کولیک، دئوکسی کولیک، کنودئوکسی کولیک و لیتوکولیک اسید)، لسیتین (مخلوطی از فسفولیپیدهای آمفیفیلیک) و ترکیبات مشتق از کلسترول است. این ترکیب دوگانه دوست، امولسیفایری است که از یک طرف به مولکولهای چربی و از طرف دیگر به مولکولهای آب متصل میشود. تشکیل امولسیون سبب محلول شدن مولکولهای چربی و افزایش مساحت سطح دردسترس آنزیم لیپاز پانکراس خواهد شد.

آنزیم لیپاز پانکراس، دیآسیل گلیسرولهای ورودی از معده را به مونوگلیسیرید و یک اسید چرب تجزیه میکند. برای انتقال این مولکولها به سیتوپلاسم سلولهای جذبکننده در میکروویلی، نیاز به تشکیل میسل به کمک صفرا است. در سیتوپلاسم سلول تریگلیسرید دوباره تشکیل میشود و همراه کلسترول، فسفولیپید و پروتئینها ساختاری به نام کیلومیکرون تشکیل میدهد که چربیها را به سیستم لنفاوی و سپس از راه سیاهرگ زیرگلویی به خون انتقال میدهند. جذب کلسترول در لوله گوارش بسیار کمتر از تریگلیسیریدها و فسفولیپیدها است . مصرف مواد غذایی حاوی فیبر (پلیساکارید سلولز) ازجمله سبزیجات و میوههای تازه، با اتصال به صفرا و کلسترول از جذب آنها در روده کوچک جلوگیری میکند.
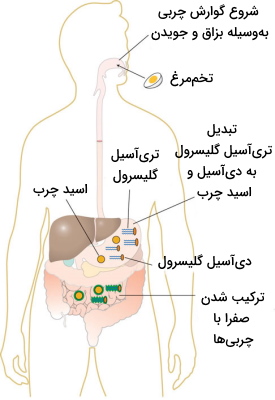
تنظیم هورمونی گوارش در روده باریک
گوارش غذا در روده وابسته به تحریک دستگاه عصبی و هورمونهایی است که حرکات این بخش از لوله گوارش و ترشح آنزیمها و صفرا را کنترل میکنند. سکرتین یکی از هورمونهای پپتیدی دستگاه گوارش است که در سلولهای S موجود در غده لیبرکان ترشح میشود. اتصال این هورمون به رسپتور آن در سطح سلولهای اگزوکرین پانکراس ترشح شیره لوزالمعده را تحریک میکند. کولهسیستوکینین (CCK) یکی دیگر از پپتیدهای مهم در تنظیم هورمونی دستگاه گوارش است. ژن این هورمون در سلولهای I غدد لیبرکان قرار دارد و پس از رونویسی و ترجمه، هورمون را به خون ترشح میکند. اتصال این هورمون به گیرندههای آن در سطح سلولهای اگزوکرین لوزالمعده و کبد ترشح شیره پانکراس و صفرا را از مجرای مشترک به دئودئوم روده تحریک میکند.
به علاوه ورود لیپیدها و اسیدهای چرب به دئودئوم سبب ترشح هورمون انتروگاسترون میشود. این هورمون با کاهش حرکات روده و معده فرصت کافی برای فعالیت لیپاز و هیدرولیز چربیها فراهم میکند. اختلال در ترشح شیره پانکراس منجر به افزایش پروتئینها و مولکولهای چربی گوارش نشده در مدفوع و ایجادد بیماری مدفوع چرب یا «استئوریا» (Steatorrhea) میشود.
گوارش در روده بزرگ
روده بزرگ بخش انتهایی لوله گوارش است که ساختار و بافتی شبیه روده کوچک دارد. اما تفاوتهای قابل توجهی بین اپیتلیال دیواره این دو اندام وجود دارد. برای مثال تاخوردگیهای حلقوی و ویلی در روده بزرگ وجود ندارند. به جز کانال مقعدی، سلولهای پوششی مخاط روده بزرگ، سلولهای استوانهای سادهای هستند که بیشتر از سلولهای جامی (گابلت) و انتروسیتها (سلولهای جذبی) تشکیل میشوند. به علاوه دیواره روده بزرگ حفرههای بیشتری تشکیل شده است که محل قرارگیری سلولهای جامی و جذبی هستند. سلولهای جامی وظیفه تولید و ترشح مخاط معده را برعهده دارند که حرکت مواد در این اندام را تسهیل و از سلولها در برابر اسید تولید شده بهوسیله باکتریها محافظت میکند. انتروسیتها آب و الکترولیت را جذب میکنند.
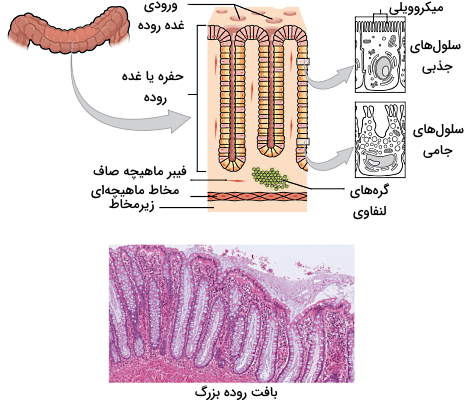
برخلاف دیگر بخشهای لوله گوارش، هیچ آنزیم گوارشی در روده بزرگ وجود ندارد و تنها گوارش شیمیایی در این اندام، تخمیر سلولز بهوسیله باکتریهای کولون است. به همین دلیل میتوان گفت بیشتر نقش روده بزرگ در لوله گوارش، بازجذب آب و یونها و تشکیل مدفوع است. سکوم، کولون، رکتوم و مجرای مقعد قسمتهای محتلف روده بزرگ در انسان هستند.
- سکوم: شش سانتیمتر ابتدایی روده بزرگ سکوم نام دارد که بهوسیله اسفنکتر ایلئوسکرال با روده کوچک در ارتباط است و آب و یونهای موجود در کیموس را جذب میکند. آپاندیس زائده کوچکی است که به این بخش از روده بزرگ متصل میشود.
- کولون: کولون بخش بعدی روده بزرگ است که از بخشهای مختلفی تشکیل میشود و باکتریهای تولیدکننده ویتامین k که سلولز را هیدرولیز میکنند در این بخش قرار دارند. تخمیر سلول بهوسیله این باکتریها با تولید گاز هیدروژن، دیاکسید کربن و متان همراه است.
- رکتوم: رکتوم بخش انتهایی روده بزرگ (حدود ۲۸٫۳ سانتیمتر) است.
- مجرای مقعد: باقیمانده گوارش نشده غذا وارد مجرای مقعد میشود. دو اسفنکتر در اینتهای این بخش از لوله گوارش وجود دارد. اسفنکتر داخلی از ماهیچههای صافی تشکیل شده و باز شدن آن غیراردی است. اما ماهیچه خارجی از ماهیچههای مخطط تشکیل میشود که تحت کنترل فرد است.

گوارش مکانیکی در روده بزرگ
گوارش مکانیکی غذا در روده بزرگ با ورود کیموس به سکوم و تحریک گیرندههای کششی شروع میشود. سه نوع انقباض ماهیچههای صاف دیواره، گوارش مکانیکی در این بخش از لوله گوارش را هدایت میکنند. حرکات قطعهقطعه کننده اولین حرکاتی هستند که با ورود غذا به این بخش شروع میشوند. در بخش قبلی توضیح دادیم که این حرکات تنها در روده بزرگ و کوچک ایجاد میشوند و در روده بزرگ به فشرده تر شدن باقیمانده غذا و بازجذب آب از سلولهای دیواره کمک میکنند.
دومین حرکت روده بزرگ، حرکات پریستالتیک یا دودی است که در تمام بخشهای این لوله از مری تا روده ایجاد میشود. این حرکات در روده بزرگ بسیار آهستهتر از بخشهای دیگر است. انقباض بزرگ یکی دیگر از حرکاتی است که تنها در این بخش از لوله گوارش ایجاد میشود. این حرکات در میانه بخش عرضی کولون شروع میشود و کیموس را سریع به رکتوم منتقل میکند.
جذب آب در روده
بیشتر آب و یونها در روده کوچک و بخش کمی از آن در روده بزرگ جذب میشود. جذب آب فشار اسمزی سلولها را تغییر میدهد و به همین دلیل وابسته به یونهای سدیم و کلر است. با همانتقالی آمینواسیدها و گلوکز وارد سلولهای دیواره روده و بهوسیله پمپ سدیم-پتاسیم غشای بازولترال وارد فضای میان بافتی میشود. خروج سدیم از سلول با افزایش فشار اسمزی و ورود آب به انتروسیتها همراه است.
سوالات متداول
در این بخش به تعدادی از سوالات پیرامون گوارش در روده پاسخ میدهیم.
گوارش کدام مواد در روده انجام می شود ؟
گوارش پروتئینها، دیساکاریدها و چربیها در روده کوچک انجام میشود. اما روده بزرگ هیچ آنزیم گوارشی ندارد و بیشتر در فشرده کردن مواد برای دفع و جذب آب و الکترولیت شرکت میکند. از آنجایی که سلولاز در لوله گوارش انسان وجود ندارد، فیبرهای سلولزی موجود در مواد غذایی گیاهی بهوسیله باکتریهای موجود در روده بزرگ تخمیر میشوند.
غذای گوارش یافته چگونه به روده بزرگ منتقل می شود ؟
حرکات دودی یا پریستالتیک، انقباضهای غیرارادی ماهیچه صاف دیواره روده باریک هستند که انقباض منظم آنها کیموس یا غذای گوارشیافته را از انتهای ژژنوم و پس از عبور از اسفنکتر ایلئوسکال به سکوم روده بزرگ منتقل میکند.
جمع بندی
در این مطلب گوارش مکانیکی و شیمیایی در روده بزرگ و کوچک را توضیح دادیم. گوارش مکانیکی مواد غذایی از دهان شروع میشود و بهوسیله حرکات معده، روده کوچک و روده بزرگ ادامه مییابد. همچنین، گوارش شیمیایی پلیمرهای زیستی غذا، بهوسیله آنزیمهای موجود در بخشهای مختلف لوله گوارش انجام میشود. گوارش کربوهیدراتها و چربیها بهوسیله آمیلاز بزاق و لیپاز زبانی در دهان شروع میشود. گوارش کربوهیدرات بهوسیله دیساکاریدازهای حاشیه مسواکی و چربیها بهوسیله لیپاز پانکراس تکمیل میشود. اما گوارش پروتئینها بهوسیله اسید و پپسین معده شروع و به کمک دیپپتیدازهای غشای میکروویلی در روده باریک پایان مییابد.









