



انواع نورون چیست؟ – به زبان ساده + شکل و عملکرد
نورونها سلولهای بسیار تخصصی و تمایزیافته بدن جانوران هستند که بین اندامها مختلف بدن و مغز که مرکز اصلی کنترل بسیاری از اعمال بدن است، ارتباط برقرار میکنند. با وجود اینکه شکل تمام این سلولها و وظیفه آنها یکسان است، بر اساس تعداد زوائد سیتوپلاسمی، سرعت انتقال جریان الکتریکی و جهت انتقال جریان الکتریکی این سلولها را به انواع مختلف تقسیم میکنیم. در این مطلب شکل انواع نورون و تقسیمبندی ان را توضیح میدهیم.
- با اجزای نورون آشنا میشوید و وظیفه هر قسمت را یاد خواهید گرفت.
- میآموزید نورونهای حسی، حرکتی و بینابینی چگونه عمل میکنند.
- با ساختار و ویژگیهای انواع نورون تکقطبی، دوقطبی و چندقطبی آشنا خواهید شد.
- میآموزید انتقال پیام در نورونهای میلیندار و بدون میلین چه فرقی دارد.
- سیناپسهای شیمیایی و الکتریکی و نحوه انتقال پیام را یاد میگیرید.
- یاد خواهید گرفت کانالهای یونی و گره رانویه چه نقشی در انتقال عصبی دارند.
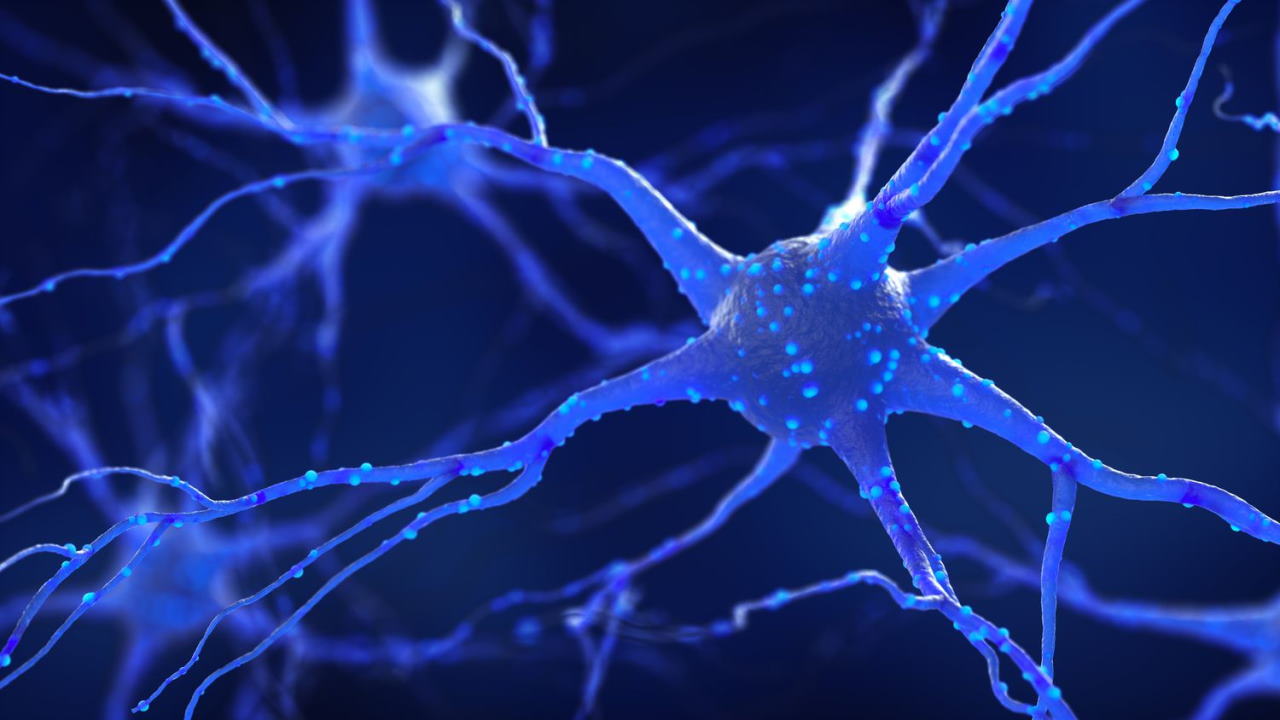


نورون چیست؟
نورونها سلولهای جریانساز و تحریک پذیر بدن انسان هستند که سیستم عصبی را میسازند. تمایز در این سلولها با از بین رفتن توانایی تقسیم میتوز و تغییر مورفولوژی سلولها همراه شده است. هر نورون از سه قسمت اصلی دندریت، جسم سلولی و آکسون تشکیل میشود.
- جسم سلولی: جسم سلولی، سوما یا جسم یاختهای هر نورون بیشترین حجم سیتوپلاسم را به خود اختصاص میدهد و به دلیل وجود هسته و شبکه اندوپلاسمی، واحد سنتزی انواع نورون به حساب میآید. شکل این بخش در بیشتر نورونها دایرهای است. تمام انتقالدهندههای عصبی پروتئینی در این بخش نورون سنتز میشود. اندازه جسم سلولی بین ۳ تا ۱۸ میلیمتر در انواع نورون متفاوت است.
- دندریت: دندریتها زوائد سیتوپلاسمی جسم سلولی هستند که پیام عصبی را از پایانه آکسون نورونهای دیگر دریافت میکنند. تعداد شاخههای دندریتی در انواع نورون متفاوت است. خار دندریتی برآمدگیهای کوچک غشای پلاسمایی دندریت است که به دلیل آرایش متفاوت اسکلت سلولی این بخش از نورون ایجاد میشود و در تشکیل سیناپس شرکت میکند.
- آکسون: آکسون رشتهای سیتوپلاسمی بلندی است که اندازه آن در نورونهای مختلف به صد برابر قطر جسم سلولی میرسد و بخش اصلی انتقال جریان الکتریکی در انواع نورون است. این بخش نورون از هیلاک، رشته آکسونی و پایانه آکسون تشکیل شده است.
- هیلاک: هیلاک قسمت ابتدایی آکسون است که از جسم سلولی خارج میشود. تعداد زیادی کانال ولتاژی سدیم در غشای پلاسمایی این بخش از نورون وجود دارد و در شروع پتانسیل عمل نقش مهمی دارد. پتانسیل عمل ایجاد شده در دندریت و جسم سلولی در پیش از انتقال به فیبر آکسونی در این ناحیه تجمیع میشود.
- رشته آکسونی: رشته آکسون بخش بلندی بین هیلاک و پایانه است که از غشای پلاسمایی یا کانالهای ولتاژی فراوان، رشتههای اکتین اسکلت سلولی برای انتقال وزیکولهای سیناپسی بین پایانه و جسم سلولی و سیتوپلاسم تشکیل میشود. این بخش در انواع نورون میلیندار یا بدون میلین است.
- پایانه آکسون: پایانه آکسون از نظر جایگاه آناتومیک و عملکرد نقطه مقابل هیلاک است. این بخش از نورون در انتهای رشته آکسونی قرار دارد و با آزاد کردن انتقالدهندههای عصبی پتانسیل عمل را به سلولهای پسسیناپسی منتقل میکند.

ساختار و عملکرد پایه تمام نورونها یکسان است. تمام این سلولها از سه بخش تشکیل شدهاند و جریان الکتریکی را منتقل میکنند. اما تفاوت آنها در محل قرارگیری سه بخش سلولی، تعداد زوائد سیتوپلاسمی، نوع انتقالدهنده عصبی که در انتقال جریان الکتریکی نقش دارد و جهت انتقال جریان بین سیستم عصبی مرکزی و محیطی است.
انواع نورون از نظر عملکرد
نورونها از نظر عملکرد به سه گروه حسی، حرکتی و بینابینی تقسیم میشوند. شکل نورونهایی که عملکرد یکسانی دارند شبیه به هم است. نورونهای حسی در پاسخ به تحریکهای لمسی، دمایی، تغییر غلظت شیمیایی مواد، تغییر فشار محیط یا امواج صوتی فعال میشوند و جریان الکتریکی را از اندامهای محیطی به دستگاه عصبی مرکزی منتقل میکنند. نورونهای حرکتی جریان الکتریکی را از بخشهای فرماندهنده سیستم عصبی مرکزی به اندامهای محیطی (ماهیچه یا غدد) منتقل میکنند. نورونهای بینابینی سلولهای عصبی کوتاهی هستند که بین نورونهای حسی و حرکتی در نخاع یا مغز ارتباط برقرار میکنند. نورونهای حسی و حرکتی در مجموعهای به نام عصب کنار هم قرار میگیرند و همزمان پیام عصبی را بین اندامهای محیطی و بخشهای مرکزی منتقل میکنند.
نورون های حسی
نورونهای حسی یا آوران بخشی از سیستم عصبی محیطی بدن هستند که یا انتهای دندریتی آنها گیرنده حسی است (گیرندههای حس درد) یا با سلولهای تحریکپذیر گیرنده (گیرندههای چشایی زبان) در ارتباط است. جسم سلولی این نورونها در گانگلیون ریشه پشتی نخاع قرا دارد و آکسون آنها پیام عصبی را به نورونهای نخاع منتقل میکند. محرک این نورونها ممکن است عوامل خارج از بدن ازجمله تغییر دما، امواج صوتی، مزه یا بوی غذا، نور خورشید و نیروی مکانیک یا محرکهای داخلی ازجمله تغییر فشار هیدروستاتیک، کشش ماهیچه اسکلتی و گازهای خون باشد. گیرندههای بویایی، شنوایی، بینایی، چشایی و مکانیکی به محرکهای خارجی و گیرندههای درد، دما و شیمیایی به محرکهای خارجی و داخلی پاسخ میدهند.
- گیرندههای بویایی: گیرندههای بویایی یکی از انواع گیرندههای شیمیایی هستند که پتانسیل غشای آنها در پاسخ به مولکول بوی محلول در مخاط بینی، تغییر میکند. این گیرندهها از انواع نورونهای دوقطبی هستند.
- گیرندههای شنوایی: گیرندههای شنوایی یا سلولهای مویی سلولهای تمایزیافته گوش داخلی هستند که پتانسیل غشای آنها در اثر لرزش امواج صوتی تغییر میکند. این سلولها با نورونهای حسی در ارتباط هستند که پیام عصبی را به مراکز کنترلی مغز انتقال میدهد.
- گیرندههای بینایی: گیرندههای بینایی سلولهای تحریکپذیر تخصصیافتهای در شبکیه چشم هستند که نورونهای حسی تحریک پتانسیل الکتریکی آنها را به بخشهای کنترلی مغز منتقل میکند.
- گیرندههای چشایی: ساختار گیرندههای چشایی بسیار شبیه گیرندههای بویایی است. سلولهای چشایی از انواع سلولهای تحریکپذیری در جوانههای چشایی زبان هستند که تغییر پتانسیل الکتریکی آنها بهوسیله نورونهای حسی به بخشهای کنترلی مغز منتقل میشود.
- گیرندههای مکانیکی: این گیرندههای محیطی انتهای نورونهای حسی هستند که در برای کاهش آستانه تحریکپذیری کپسولی از بافت پیوندی اطراف آنها قرار دارد. کشیدگی یا فشردگی کپسول اطراف این نورونها با ایجاد جریان الکتریکی و انتقال پیام عصبی به بخشهای کنترلی سیستم عصبی مرکزی همراه است.
- گیرندههای حس وضعیت: این گیرندهها نورونهای حسی عضلات اسکلتی و از انواع گیرندههای مکانیکی هستند. کشش ماهیچه وضعیت کانالهای یونی این نورونهای حسی را و پتانسیل الکتریکی آن را تغییر میدهد.
- گیرندههای درد یا نوسیسپتورها (Nociceptors): این نورونهای حسی یکی از انواع گیرندههای مکانیکی هستند که تغییرات دمای یا فشار (درد) محیط را شناسایی و برای جلوگیری از آسیب بافتها به بخشهای کنترلی مغز پیام میفرستند. این نورونها در لایههای دور از سطح پوست و اندامهای داخلی قرار دارند.
- گیرندههای شیمیایی: گیرندههای شیمیایی در دیواره رگهای محیطی و مغز قرار دارند و در پاسخ به تغییرات شیمیایی خون ازجمله کاهش فشار جزئی اکسیژن در خون تحریک میشوند. اجسام آئورتی و کاروتیدی گیرندههای شیمیایی و سلولهای تمایزیافتهای هستند که پتانسیل الکتریکی ایجاد شده در آنها به نورون حسی و سپس دستگاه عصبی مرکزی منتقل میشود.

نورورنهای حسی ناحیه سر و گردن بخش حسی اعصاب جمجمهای (۱۲ جفت) یا «کرانیال» (Cranial Nerves) را میسازند و پیام عصبی (به جز گیرندههای بویایی) را ابتدا به ساقه مغز و سپس بخشهای بالایی مغز منتقل میکنند. نورونهای حسی اندامهای پایین گردن بخش حسی اعصاب نخاعی (۳۱ جفت) را تشکیل میدهند که پیام عصبی آنها ابتدا به نخاع سپس بصلالنخاع، تالاموس و بخشهای کنترلی مغز منتقل میکنند.
نورون های حرکتی
نورونهای حرکتی انواعی از این سلولهای عصبی هستند که آکسون بلند آنها پیام بخشهای کنترلی سیستم عصبی مرکزی را به ماهیچهها و غدد اگزوکرین منتقل و با ایجاد پتانسیل عمل این بافتها را تحریک میکنند. جسم سلولی این نورونها در قشر حرکتی مخ، ساقه مغز یا نخاع قرار دارد. نورونهای حرکتی را میتوان بر اساس جایگاه آناتومی به دو دسته بالایی و پایینی تقسیم کرد.
نورونهای حرکتی بالایی
دندریت و جسم سلولی این نورونهای مخروطی در قشر حرکتی مخ قرار دارد و آکسون این نورونها در شاخ شکمی نخاع با نورونهای بینابینی یا مستقیم با نورونهای حرکتی پایینی سیناپس تشکیل میدهد.
نورونهای حرکتی پایینی
دندریت نورونهای حرکتی پایینی در نخاع قرار دارد و آکسون آنها پیام عصبی را به اندامهای داخلی و سطحی بدن منتقل میکند. این نورونها، رشتههای وابران دستگاه عصبی هستند. این نورونها به سه دسته نورونهای حرکتی سوماتیک، نورونهای حرکتی احشای ویژه (صورت) و نورونهای حرکتی احشای عمومی (اندامهای داخلی) تقسیم میشوند. نورونهای حرکتی احشای با ماهیچههای تغییر حالت چهره، جویدن، تکلم و بلع سیناپس تشکیل میدهند. این نورونها بخش حرکتی اعصاب جمجمهای چشمی-حرکتی، دورکننده، قرقرهای و عصب زیرزبانی را تشکیل میدهند.
این نورونهای حرکتی پایینی با رشتههای عصبی خودمختار (سمپاتیک و پاراسمپاتیک) در گانگلیونهای نخاعی سیناپس تشکیل میدهند و به طور غیرمستقیم انقباض ماهیچههای صاف لوله گوارش، دیواره رگها، ماهیچه قلبی، ماهیچه صاف دیواره مجاری ادراری و مثانه را تحریک یا مهار میکنند. نورونهای حرکتی سوماتیک آلفا، بتا و گاما با ماهیچههای اسکلتی سیناپس میدهند.
- نورونهای حرکتی آلفا: جسم سلولی این نورونها در شاخ شکمی نخاع قرار دارد و پایانه آکسون هر کدام از آنها به طور میانگین با ۱۵۰ فیبر ماهیچه اسکلتی سیناپس میدهد. به مجموع نورون حرکتی و تمام فیبرهای ماهیچههای آن یک واحد حرکتی گفته میشود که بر اساس سرعت تحریک الکتریکی به سه دسته آهسته، سریع و مقاوم به خستگی تقسیمبندی میشوند.
- واحد حرکتی آهسته: فیبر عصبی این واحد حرکتی با تعداد کمی افیبرهای ماهیچه اسکلتی سیناپس دارد و بخش بسیار کمی از نیروی انقباض را فراهم میکند اما نسبت به خشتگی بسیار مقاوم است. به همین دلیل این واحد حرکتی در حرکاتی که به انقباض پیوسته نیاز است مثل (صاف نگهداشتن پشت) فعال میشود. ماهیچههای این واحد حرکتی بخش اصلی انرژی خود را از واکنشهای اکسیداسیون میتوکندری تامین میکنند و به دلیل میوگلوبین فراوان به آنها فیبر ماهیچهای قرمز نیز گفته میشود.
- واحد حرکتی سریع: نورونهای حرکتی این واحد حرکتی با فیبرهای ماهیچهای زیادی سیناپس دارند. به همین دلیل بخش بیشتری از نیروی انقباضی ماهیچه را فراهم میکنند، اما به سرعت خسته میشوند. این واحدها در حرکاتی که نیاز به نیرو و انرژی زیاد در کوتاهمدت دارد (مثل پریدن یا دویدن) فعال میشوند. بیشتر انرژی فیبرهای ماهیچهای این واحدهای حرکتی از مسیر گلیکولیز تامین میشود و به دلیل تجمع کمتر میوگلوبین در سارکوپلاسم به آنها فیبرهای سفید نیز گفته میشود.
- واحد حرکتی مقاوم به خستگی: تعداد فیبرهای ماهیچهای این واحدهای حرکتی بین واحدهای حرکتی آهسته و سریع است. سرعت انقباض ماهیچههای این واحد حرکتی (تحریک جریان الکتریکی) کمتر از فیبرهای سریع اما مدت زمان منقبض ماندن ماهیچه بیشتر از آنها است. نیروی ایجاد شده از انقباض فیبر ماهیچه واحد حرکتی مقاوم به خستگی از فیبرهای سریع بیشتر است و از مسیر اکسیداسیون و گلیکولیز انرژی مورد نیاز خود را تامین میکند.
- نورونهای حرکتی بتا: آکسون نورونهای حرکتی بتا با سلولهای دوک ماهیچه سیناپس میدهد. این سلولها گیرنده حس موقعیت هستند که در اثر کشش بیش از تحمل ماهیچه تحریک میشوند.
- نورونهای حرکتی گاما: نورونهای حرکتی گاما با سلولهای دوک ماهیچه سیناپس میدهند و حساسیت این سلولها به کشش ماهیچه را تنظیم میکنند.
نورون های بینابینی
نورونهای بینابینی در سیستم عصبی مرکزی بین نورونهای حسی و حرکتی ارتباط برقرار میکنند. این نورونها بین ۲۰ تا ۳۰٪ کل نورونهای نئوکوتکس مخ را به خود اختصاص میدهند و وظیفه آنها برقراری ارتباط (انتقال جریان الکتریکی) بین لوبهای مختلف مخ است. این نورونها به دو دسته منطقهای و تقویتی تقسیم میشوند.
- نورونهای بینابینی منطقهای: این نورونها آکسونهای کوتاهی دارند که در تشکیل مدارهای کوچک در یک منطقه مغز برای پردازش اطلاعات کم ارتباط برقرار میکنند.
- نورونهای بینابینی تقویتی: این نورونها آکسونهای بلندی دارند که بین نورونهای دو منطقه مغز ارتباط برقرار میکنند.
بیشتر نورونهای بینابینی سیستم عصبی مرکزی، نورونهای مهاری هستند که با رهایش انتقالدهنده عصبی GABA یا گلایسین پتانسیل الکتریکی نورونها را تغییر میدهند. تعداد کمی از این نورونها از انواع تحریکی هستند که برخلاف سایر نورونهای تحریکی با رهایش انتقالدهنده عصبی گلوتامات پیام عصبی را بین نورونها منتقل میکنند.
انواع نورون از نظر شکل
با وجود ساختار پایه یکسان نورونها این سلولهای عصبی بر اساس محل قرارگیری جسم سلولی، تعداد آکسون و تعداد دندریت به چهار دسته پایه نورونهای تکقطبی، دوقطبی، چندقطبی و تکقطبی کاذب تقسیم میشود.
- نورونهای تکقطبی: در این سلولها یک زائده سیتوپلاسمی از جسم سلولی خارج میشود. این نورونها در مهرهداران وجود ندارد اما در سیستم عصبی حشرات پیام عصبی مغز را به ماهیچهها و غدد منتقل میکند.
- نورونهای دوقطبی: در این سلولها دو زائده سیتوپلاسمی (یک دندریت و یک آکسون) از جسم سلولی خارج میشود. پتانسیل الکتریکی ایجاد شده در شبکیه چشم (رتینا) به این نورونها منتقل میشود.
- نورونهای چندقطبی: بیشتر نورونهای بافت عصبی انسان از نوع چند قطبی است. در این سلولها یک زائده آکسون بلند و چند زائده دندریتی از جسم سلولی خارج میشود. سلولهای پورکنژ در درخت زندگی مخچه و سلولهای مخروطی قشر مخ دو نوع نورون چندقطبی در سیستم عصبی مرکزی هستند.
- نورونهای تکقطبی کاذب: شکل نورونهای تکقطبی کاذب به نورونهای تکقطبی و دوقطبی شباهت دارد. در این سلولهای عصبی مثل نورونهای تکقطبی یک زائده سیتوپلاسمی از جسم سلولی خارج شده که سپس مثل سلولهای دوقطبی به یک شاخه دندریتی و یک آکسون تقسیم میشود.

سلول های پورکنژ
سلولهای پورکنژ از انواع نورون های دوقطبی و مهاری مخچه هستند که بهوسیله رهایش انتقالدهنده عصبی GABA با نورونهای دیگر پسسیناپسی ارتباط برقرار میکنند. این نورونها پس از سلولهای بتز (Betz cells) بزرگترین سلولهای مغزی هستند که از یک آکسون و تعداد بسیار زیادی دندریت از جسم سلولی آنها خارج میشود. هر سلول پورکنژ با حدود ۵۰۰ نورون دیگر سیناپس تشکیل میدهد.

سلول های مخروطی
سلولهای مخروطی گروه دیگر از انواع نورون های دوقطبی هستند که در تشکیل بافت عصبی قشر مخ، هیپوکامپ و آمیگدالا (هستههای بادامی لوب پیشانی که در حافظه، تصمیمگیری و پاسخهای احساسی نقش دارد) شرکت میکنند. این نورونها سلولهای اولیه مسیر عصبی قشر مخ-نخاع هستند و پیام عصبی آنها بهوسیله نورونهای حرکتی بالای به نخاع منتقل میشود. جسم سلولی (با طول ۲۰۰ میکرومتر) این نورونها برخلاف سایر نورونهای چندقطبی مخروطی است. یک زائده آکسونی از سوما خارج میشود و دندریتها به دو بخش راسی و پایه تقسیم میشوند.
- دندریت راسی: دندریت راسی یک زائده سیتوپلاسمی بلند و ضخیم است که از راس مخروط جسم سلولی (بخشهای داخلی مغز) به قشر مخ کشیده میشود.
- دندریت پایه: سه تا پنج زائده دندریتی از بخش قاعدهای جسم سلولی خارج شده که انتهای دور آنها به شاخههای کوچکتر تقسیم میشود.

انواع سیناپس نورون
سیناپس محل برقراری دو نورون در سیستم عصبی است. این ساختار آناتومی از پایانه آکسون نورون پیشسیناپسی، فضای سیناپسی (بین دو نورون یا نورون با سلولهای غیرعصبی) و سلول پسسیناپسی (نورون یا سلولهای غیرعصبی) تشکیل شده است. سیناپس الکتریکی و شیمیایی دو نوع ساختار آناتومی و محل انتقال پیامهای عصبی بین سلولها هستند.

سیناپس الکتریکی
در سیناپس الکتریکی غشای پلاسمایی دو نورون بسیار به هم نزدیک و بهوسیله اتصالات شکافدار با هم در ارتباط هستند. به همین دلیل انتقال جریان الکتریکی در این سیناپسها سریعتر از سیناپسهای شیمیایی انجام میشود. انتقال کاتیونها از سلول پیشسیناپسی به پسسیناپسی با انتقال جریان الکتریکی همراه است. این سیناپس فقط در بافت عصبی ایجاد میشود.

سیناپس شیمیایی
در سیناپس شیمیایی جریان الکتریکی بهوسیله اتصال مولکولهای شیمیایی آزاد شده از پایانه آکسون نورونهای پیشسیناپسی به گیرندههای غشایی سلولهای پسسیناپسی منتقل میشود. این سیناپس بین دو نورونهای بافت عصبی و نورون-ماهیچه یا نورون-غدد اگزوکرین ارتباط برقرار میکند. زمانی که پتانسیل عمل به پایانه آکسون نورون پیشسیناپسی میرسد، کانالهای دریچهدار کلسیمی حساس به ولتاژ را باز میکند. باز شدن این کانال با ورود یونهای کلسیم به پایانه آکسون، اتصال آن به پروتئینهای مسیر اگزوسیتوز، اتصال وزیکولهای حاوی انتقالدهنده عصبی به غشای سلول پیشسیناپسی و خروج انتقالدهنده عصبی از پایانه آکسون همراه است.
گیرنده انتقالدهده عصبی در نورونهای پسسیناپسی کانالهای یونی دریچهداری هستند و اتصال لیگاند-گیرنده در آنها با تحریک یا مهار نورن پسسیناپسی همراه است. بر همین اساس نورونها را به انواع تحریکی و مهاری تقسیم میکنند.
- نورونهای تحریکی: اتصال انتقالدهنده عصبی آزاد شده از این نورونها به کانال دریچهدار کاتیونی در سلول پسسیناپسی با افزایش پتانسیل الکتریکی سلول پسسیناپسی و ایجاد پتانسیل عمل همراه است. آزاد شدن استیل کولین از پایانه آکسونی نورونهای حرکتی و اتصال آن به کانالهای دریچهدار سدیمی یکی از سیناپسهای تحریکی است. نورونهای تحریکی در سلول پسسیناپسی «پتانسیل پسسیناپسی تحریکی» ( Excitatory Postsynaptic Potential | EPSP) ایجاد میکنند.

- نورونهای مهاری: اتصال انتقالدهنده عصبی آزاد شده از این نورونها به کانالهای آنیونی در غشای نورونهای پسسیناپسی با منفی شدن پتانسیل الکتریکی سلول پسسیناپسی (هایپرپلاریزاسیون) و مهار سلول همراه است. پتانسیل الکتریکی ایجاد شده در سلول پسسیناپسی «پتانسیل پسسیناپسی مهاری» ( Inhibitory Postsynaptic Potentials | IPSP) نام دارد. برای مثال انتقالدهنده عصبی GABA (گاما بوتیریک اسید) به کانالهای دریچهدار کلر در غشای پلاسمایی سلول پسسیناپسی با ورود یون کلر به سلول، هایپرپلاریزاسیون و مهار سلول همراه است.
انتقالدهنده عصبی پس از ایجاد تغییر در سلول پسسیناپسی باید از فضای سیناپسی خارج و سلول برای تحریک یا مهار بعدی آماده شود. این فرایند بهوسیله آنزیمهای تجزیهکننده انتقالدهنده عصبی یا بهوسیله بازجذب آن در سلول پیشسیناپسی انجام میشود. برای مثال استیلکولین استراز آنزیمی است که در سیناپس عصب-عضله اسکلتی با تجزیه استیلکولین به تحریک عضله پایان میدهد. نورونها را بر اساس نوع انتقالدهنده عصبی آنها میتوان به انواع کولینرژیک، سرونرژیک، گابانرژیک، آدرنرژیک، دوپامینرژیک، نورآدرنرژیک و گلوتاماترژیک تقسیم میشوند. عملکرد هر یک از این نورونها در جدول زیر خلاصه شده است.
| نوع نورون | عملکرد | محل فعالیت | انتقالدهنده عصبی |
| کولینرژیک | کنترل ماهیچه، حافظه | سیستم عصبی مرکزی و محیطی | استیلکولین |
| سرونرژیک | حرکات روده، تنظیم خلق و چرخه خواب |
معده، روده و سیستم عصبی مرکزی | سروتونین |
| گابانرژیک | مهار سیستم عصبی مرکزی | مغز | GABA |
| آدرنرژیک | واکنشهای جنگ یا گریز | مدولای غده فوق کلیه | اپینفرین |
| دوپامینرژیک | حرکت ارادی ماهیچهها، عملکرد شناختی و مسیر پاداش | هیپوتالاموس | دوپامین |
| نورآدرنرژیک | واکنشهای جنگ یا گریز | مدولای غده فوق کلیه | نوراپینفرین |
| گلوتاماترژیک | تحریک نورونها، حافظه | سیستم عصبی مرکزی و محیطی | گلوتامات |
انواع نورون از نظر میلین
میلین پوششی از جنس لیپید اطراف آکسون نورونها است (دندریت و جسم سلولی میلین ندارد) که از محافظت میکند و سرعت انتقال پتانسیل عمل در این سلولها را افزایش میدهد. اما در تمام نورونها وجود ندارد. سلولهای پشتیبان سیستم عصبی مرکزی و محیطی با دو مکانیسم متفاوت میلین نورونها را تشکیل میدهند.
- نورونهای بدون میلین بخشهای خاکستری و نورونهای میلیندار بخشهای سفید بافت عصبی را تشکیل میدهند.
- قطر آکسون نورونهای بدون میلین از نورونهای میلیندار کمتر است تا کاهش سرعت ایجاد شده به دلیل بدون میلین را جبران کند.
- فیبرهای حسی C مهمترین انواع نورون بدون میلین بافت عصبی محیطی هستند که پیام گیرندههای حس لامسه (سوزش و خارش) و دما را به نخاع منتقل میکنند.
- نورونهای حرکتی بالایی و نورونهای حرکتی سوماتیک از انواع نورون میلیندار، اما نورونهای حرکتی سیستم سمپاتیک و پاراسمپاتیک از انواع بدون میلین هشتند.
تشکیل میلین
سلولهای پشتیبان شوآن میلین نورونهای سیستم عصبی محیطی و سلولهای اولیگودندروسیت میلین نورونهای سیستم عصبی مرکزی را تشکیل میدهند. هر سلول شوآن در تشکیل میلین یک آکسون و هر اولیگودندروسیت در تشکیل میلین آکسون چند نورون شرکت میکند.
- سلولهای شوآن: تشکیل میلین بهوسیله سلولهای صاف شوآن با برهمکنش پروتئینهای غشای سلول شوآن و آکسون شروع میشود. سپس غشای پلاسمایی سلولهای شوآن که مثل غشای پلاسمایی سایر سلولها از تعداد زیادی مولکول لیپیدی (به ویژه گلیکوسربرولیپید، اسفنگومیلین و کلسترول) تشکیل شده است دور آکسون نورونها میپیچید. با هر دور چرخش دور آکسون سیتوپلاسم و هسته سلولها (نورولما) از مرکز آکسونی دورتر میشود. در پایان فرایند میلیندار شدن، هسته و سیتوپلاسم سلول شوآن در سطح و بافت لیپیدی غشا در تماس با آکسون است. هر سلول شوآن لایه لیپیدی حدود یک میلیمتر اطراف آکسون تشکیل میدهد.
- اولیگودندروسیتها: اولیگودندروسیتها سلولهای پشتیبان بافت عصبی مرکزی هستند که زوائد سیتوپلاسمی آنها در تشکیل غلاف میلین نورونهای مغز و نخاع شرکت میکنند. به همین دلیل در غلاف میلین نورونهای سیستم عصبی مرکزی برخلاف نورونهای سیستم عصبی مرکزی، هسته سلولهای پشتیبان وجود ندارد.

تفاوت انتقال پیام عصبی نورون میلیندار و بدون میلین
پتانسیل عمل در هر دو نورون میلیندار و بدون میلین با مکانیسم مشابه و پس از اتصال انتقالدهنده عصبی به گیرندههای غشای نورون پسسیناپسی (کانالهای لیگاندی) تشکیل میشود. باز شدن کانالهای لیگاندی و ورود یون سدیمی پتانسیل الکتریکی غشا را مثبتتر کرده و سبب باز شدن کانالهای ولتاژی سدیم میشود. اما انتقال این جریان الکتریکی از سیناپس به پایه آکسون در نورونهای میلیندار و بدون میلین متفاوت است.
کانالهای ولتاژی سدیم در تمام طول غشای آکسون نورونهای بدون میلین وجود دارد و اختلاف پتانسیل ایجاد شده در سیناپس کانالهای ولتاژی ناحیه کناری و ورود جریان یون سدیم را تحریک میکند. این توالی تا پایانه آکسون ادامه دارد. اما در نورونهای میلیندار کانالهای سدیمی ولتاژی در گرههای رانویه (فاصله بین دو قطعه از غلاف میلین) قرار دارند. به همین دلیل جریان الکتریکی ایجاد شده در سیناپس در تمام طول آکسون منتقل نمیشود و از یک گره رانویه به گره دیگر جهش دارد. در نتیجه سرعت انتقال جریان الکتریکی در نورونهای میلیندار از نورونهای بدون میلین بیشتر است.

سوالات متدوال
در این بخش به تعدادی از سوالات متداول پیرامون انواع نورون پاسخ میدهیم.
کدام کانالهای یونی در زمان دپلاریزاسیون و کدام کانالهای یونی در زمان ریپلاریزاسیون نورون باز هستند؟
کانالهای ولتاژی سدیم در زمان دپلاریزاسیون غشای نورون و کانالهای ولتاژی پتاسیم در زمان رپلاریزاسیون باز هستند.
گره رانویه چه نقشی در انتقال جریان عصبی انواع نورون دارد ؟
گره رانویه در انواع نورونهای میلیندار وجود دارد. انتقال جریان الکتریکی در این نورونها به شکل جهشی است و بیشتر از نورونهای بدون میلین همقطر است.










سیناپس الکتریکی تنها در بافت عصبی وجود ندارد بلکه در میوکارد قلب میان سلول های ماهیچه ای نیز وجود دارد .
سلام و وقت بخیر؛
مطلب بازبینی و اصلاح شد.
از دقت و همراهی شما با مجله فرادرس سپاسگزاریم.