



نفوذپذیری انتخابی غشا چیست؟ – عوامل موثر – به زبان ساده
غشاهایی که تنها موادی با ویژگیهای خاص را از خود عبور میدهند «نفوذپذیری انتخابی» (Selectively Permeability) دارند. مشخصترین مثال غشاهایی با ویژگی نفوذپذیری انتخابی، غشای پلاسمایی و اندامکهای زیستی هستند. این ويژگی غشا، بهوسیله ساختار فسفولیپیدی و پروتئینهای ویژه موجود در آن به وجود میآید. در این مطلب علاوه بر بررسی نفوذپذیری انتخابی غشا، به بررسی ساختار و عملکرد پروتئینهایی میپردازیم که به ایجاد چنین خاصیتی کمک میکنند.




نفوذپذیری انتخابی چیست ؟
نفوذپذیری انتخابی در غشای سلولی تنها به مواد موردنیاز، اجازه ورود یا خروج از سلول را میدهد و به سلول کمک میکند محتویات داخلی خود را تحت کنترل داشته باشد. انتقال مواد از این غشا به صورت فعال و با صرف ATP یا به صورت غیرفعال و بر اساس اختلاف غلظت مواد در دو طرف غشا انجام میشود.
تفاوت نفوذپذیری انتخابی و تراوایی نسبی چیست؟
غشای دارای نفوذپذیری انتخابی و «غشای نیمهتراوا» (Semipermeable Membranes)، هر دو عبور مواد از غشا را تنظیم میکنند. این دو مفهوم یکسان نیستند اما در برخی مطالب به جای هم استفاده میشوند. غشای نیمهتراوا مثل یک فیلتر عمل میکند و ذرات را بر اساس اندازه، بار الکتریکی، حلالیت یا ویژگیهای فیزیکی و شیمیایی عبور میدهد. بعضی مواد نیز با فرایندهای غیرفعال اسمز و انتشار منتقل میشوند. اما غشای دارای نفوذپذیری انتخابی، با کمک ناقل و مصرف انرژی، مواد را از یکسو به سوی دیگر، منتقل میکند.
نیمهتراوایی در غشاهای طبیعی و سنتزی حتی فیبرها نیز وجود دارد. برای مثال، تصور کنید به برای جلوگیری از ورود حشرات برای پنجره اتاق خود توری نصب کردهاید. این توری اجازه عبور هوا را میدهد ول یاز ورود پشه به اتاق شما جلوگیری میکند. این مثال سادهای از عملکرد یک غشای نیمهتراوا است. غشای داخلی تخممرغ نمونهای از غشاهای دارای نفوذپذیری انتخابی است که علاوه بر محافظت دربرابر میکروبها ورود و خروج مواد را کنترل میکند.
بهترین مثال برای غشایی است که ویژگی نفوذپذیری انتخابی و نیمهتراوایی را همزمان دارد، غشای پلاسمایی است. در ادامه به بررسی ویژگیهای مرتبط با نفوذپذیری انتخابی در این غشا میپردازیم.
ساختار غشا با نفوذپذیری انتخابی چیست ؟
غشای سلولی ساختاری دولایه از فسفولیپیدها است. سرهای آبدوست فسفولیپید به سمت خارج و داخل سلول و دو سر آبگریز برای تشکیل دولایه کنار هم قرار گرفتهاند. حضور مولکولهای زیر در غشای سلولی انسان، بین فسفولیپیدها فاصله ایجاد میکند. لازم به ذکر است این مولکولها به صورت عرض غشایی، درونغشایی یا متصل به سطح غشا قرار میگیرند.
عملکرد غشا با نفوذپذیری انتخابی چیست؟
این ویژگی غشا و مجزا نگهداشتن محیط داخلی سلول و اندامکها از محیط اطراف نقش بسیار مهمی در عملکرد برخی از سلولها و اندامکها، ازجمله سلولهای ماهیچه قلب، نورونها و میتوکندری دارد. عملکرد نورون و سلوهای ماهیچه وابسته به ایجاد اختلاف پتانسیل در غشای سلولی است و نفوذپذیری انتخابی غشا، این کار را تسهیل میکند.
نحوه عبور مواد از غشا با نفوذپذیری انتخابی
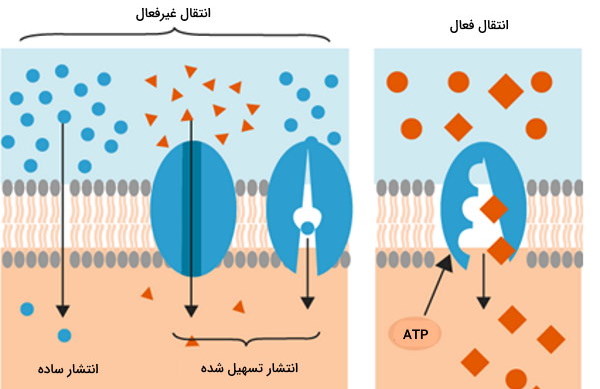
عبور مواد از غشا با نفوذپذیری انتخابی به دو صورت فعال و غیرفعال انجام میشود.
- انتقال غیرفعال: دو نوعِ انتشار ساده و انتشار تسهیل شده است.
- انتشار ساده: حرکت مواد در جهت اختلاف غلظت بدون مصرف انرژی است. انتقال پروتئینهای کوچک و بدون بار مثل اتانول، و دیگر گازها ابهوسیله این روش انجام میشود.
- انتشار تسهیل شده: حرکت مواد بزرگتر مثل گلوکز و قطبی نیاز به کمک یک پروتئین دارد. این پروتئین بدون مصرف ATP و با تغییر کنفورماسیون، ماده را از یک طرف غشا به سمت دیگر منتقل میکند. حرکت آب در آکوآپورین از این نوع است.
- انتقال فعال: انتقال مواد در این روش برخلاف گرادیان غلظت است. این انتقال به دو دسته اولیه و ثانویه تقسیم میشود.
- اولیه: این نوع انتقال نیاز به مصرف ATP یا GTP دارد. در این نوع انتقال انرژی لازم، با شکستن پیوند فسفودیاستری موجود در نوکلئوتید تامین میشود.
- ثانویه: در این حالت انرژی لازم جهت انتقال مواد در خلاف جهت گرادیان غلظت، از انرژی گرادیان الکتروشیمیایی - حاصل از انتقال فعال اولیه مواد - تامین میشود. انتقال مواد در این حالت به دو شکل همسو (Symport) و ناهمسو (Antiport) انجام میگیرد.
انتشار تسهیل شده
انتشار تسهیل شده حرکت مواد از عرض غشا بهکمک ناقل پروتئینی است که به سه روش انجام میگیرد.
- «تکانتقالی»: یک مولکول در جهت شیب غلظت منتقل میشود.
- انتقال همسو: یک یون و به همراه مولکول دیگر در خلاف جهت شیب غلظت منتقل میشود.
- انتقال ناهمسو: یک یون و یک مولکول در دو جهت و خلاف شیب غلظت منتقل میشوند.

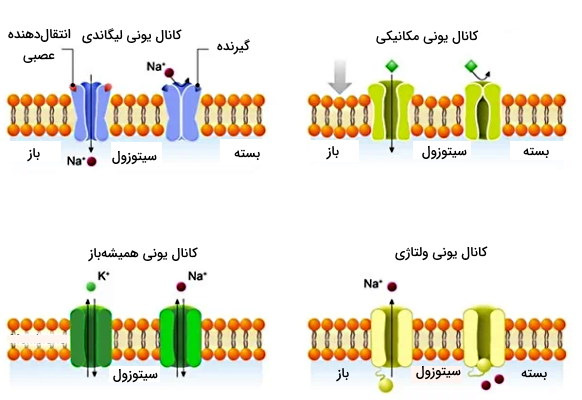
کانالهای یونی
کانالهای یونی، پروتئینهایی در غشای پلاسمایی هستند که یونهای معدنی را عبور میدهند. این پروتئینها ۴ نوع دارند که در زیر آورده شدهاند.
کانال یونی لیگاندی
این کانالها با اتصال لیگاند به دریچه باز میشوند. یک دسته از کانالهای غشای نورون از این نوع است که با اتصال انتقالدهنده عصبی به دریچه و تغییر کنفورماسیون پروتئین عرض غشایی، یون را از غشا عبور میدهد. گلوتامات و استیلکولین دو انتقالدهنده عصبی مهم در مغز هستند که باعث باز شدن کانالهای دریچهدار کاتیونی در مغز و ایجاد پتانسیل غشا در نورون میشوند. در عوض «GABA» و «گلایسین» کانالهای و را باز و ایجاد پتانسیل غشا را مهار میکنند. این کانالها هدف داروهای درمان بیماریهای مختلف ازجمله افسردگی، اضطراب، صرع، آلزایمر، شیزوفرنی و اوتیسم هستند.
کانال یونی مکانیکی
این کانالها که به عنوان یک گیرنده حسی عمل میکنند، در پاسخ به تغییرات فشار، لمس و کشش، تغییر شکل و به یونها اجازه عبور میدهند. این کانالها در سلولهای موی گوش میانی، تحریکهای مکانیکی امواج صوتی و حرکت سر را به جریان الکتریکی غشا تبدیل میکنند.
کانال یونی همیشه باز
کانالهای همیشهباز (بدون دریچه یا نشتی) یونها را براساس اختلاف غلظت عبور میدهند. بهطور مثال میتواند از طریق این کانالها بهراحتی و بر اساس شیب غلظت خارج شود.
کانال یونی وابسته به ولتاژ
مهمترین کانالهای ولتاژی، کانالهای ، و هستند. مسئولیت ایجاد پتانسیل عمل در غشای سلولهای عصبی و ماهیچهای بر عهده کانالهای و است و فعالیت آنها سبب ایجاد سیگنال عصبی در نورون و انقباض در ماهیچه میشود. باز شدن این کانالها، افزایش یون مثبت داخل سلول و قطبی شدن غشا را بهدنبال دارد.
کانالهای ولتاژی نقش مهمی در مسیرهای پیامرسانی دارند. ورودی از این کانال، پیامرسان ثانویه و آغازگر بسیاری از فرایندهای سلولی است. این کانال در انقباض ماهیچه قلبی و صاف، ترشح هورمونها، فعال شدن پروتئینکیناز، شروع انتقال سیناپسی و تنظیم بیان ژن نقش دارد.
جهش در کانالهای یونی یا بروز بیماریهای خودایمنی با هدف قرار دادن این کانالها، عملکرد آنها را دچار اختلال میکنند. پراکندگی کانالهای یونی در اندامهای مختلف، در ایجاد بسیاری از بیماریها نقش دارد.
کانالهای یونی هدف درمان
در بعضی بیماریها ممکن است علت اختلال ایجاد شده، کانالهای یونی موجود در غشا باشد. به همین دلیل داروهای تجویزی در این بیماریها، کانال را هدف اثر خود قرار میدهند. در حال حاضر تعداد کمی از این داروها استفاده میشوند که مربوط به بیماریهای قلبی، بیماریهای سیستم عصبی و بیحسکنندههای موضوعی هستند. برای نمونه «کاربامازپین» (Carbamazepine) یکی از اولین داروهای موثر شناخته شده بر کانالهای یونی است. این دارو که در درمان صرع و دیگر بیماریهای عصبی استفاده میشود، با مهار کانالهای ولتاژی سدیم در نورون، از پلاریزه شدن غشا و تحریک بدونتوقف عصب جلوگیری میکند.
«بیحسکنندههای موضعی» (Local Anesthetics) مولکولهای «دوگانهدوستی» (Amphipathic) هستند که مکانسیم عمل اصلی در آنها مهار گیرندههای حسی بهوسیله مهار کانالهای ولتاژی سدیم است. این ترکیبات، بعضی کانالهای پتاسیمی و کلسیمی را نیز مهار میکنند. لیدوکائین یکی از این داروها است عوامل زیر، بهرهگیری از این دارو را محدود میکند.
- اتصال ضعیف به کانال
- عدم عملکرد اختصاصی
ناقلهای گلوکز
گلوکز مولکولی آبدوست است که بهدلیل اندازه بزرگش نمیتواند بهوسیله انتشار ساده از عرض غشا عبور کند. به همین دلیل به ناقل غشایی نیاز دارد. ناقلهای گلوکز، از انرژی الکتروشیمیایی برای انتقال ماده استفاده میکنند و میتوان آنها را در هر دو گروه ناقلهای انتشار تسهیل شده و انتقال فعال دستهبندی کرد. فسفریله شدن گلوکز ورودی از این کانالها، اجازه نمیدهد مولکول از سلول خارج شود. دو نوع ناقل گلوکز در بدن انسان وجود دارد.
- GLUTs: چهارده نوع از این ناقل در بدن انسان وجود دارد که همه وابسته به و ATP (بهطور غیرمستقیم) هستند. این پروتئین عرض غشایی، ۱۲ بار از غشا عبور میکند.
- SGLTs: کانالهای وابسته به هستند و برای ایجاد شیب غلظت به ATP نیاز دارند. در چند اندام از جمله روده، توبولهای کلیه و سد خونی-مغزی قرار دارند.
GLUTs
نامگذاری این ناقلها براساس بخش اول نام انگلیسی آن (Glucose Transporter) (GLUT)، همراه با عدد نشاندهنده بافت و اندامهای مختلف، انجام میگیرد. این ناقلها به سه دسته کلی تقسیم میشوند.
- دسته اول (Class I): شامل GLUT1-GLUT4
- دسته دوم (Class II): شامل GLUT5 و GLUT6 و GLUT9 و GLUT11
- دسته سوم (Class III): شامل GLUT12 و GLUT10 و GLUT8 و GLUT6 و GLUT13
این ناقلها جزئی از انتشار تسهیل شده هستند و گلوکز را از خارج سلول (غلظت پایین گلوکز) به داخل سلول (غلظت بالای گلوکز) منتقل میکنند. GLUTs از انرژی گرادیان الکتروشیمیایی یون سدیم استفاده و گلوکز را بهصورت همانتقال با این یون وارد سلول میکنند. اتصال گلوکز به پروتئین باعث تغییر در کنفورماسیون آن و ورود گلوکز به سلول میشود. GLUT1-GLUT5 ناقلهایی هستند که ویژگی و عملکرد آنها بهخوبی شناخته شده است. در جدول زیر عملکرد و محل قرارگیری هر نوع را بررسی میکنیم.
| محل قرارگیری | ویژگی و عملکرد | |
| GLUT1 |
| عدم وابستگی به انسولین |
| GLUT2 |
عدم وابستگی به انسولین تمایل کم به گلوکز | |
| GLUT3 |
عدم وابستگی به انسولین تمایل زیاد به گلوکز | |
| GLUT4 |
| وابسته به انسولین |
| GLUT5 | سلولهای اپیتلیوم روده |
عدم وابستگی به انسولین انتقال فروکتوز |
تمایل پایین ناقلها به گلوکز به این معنی است که تنها در صورت غلظت بالای گلوکز، انتقال صورت میگیرد و تمایل بالا نشان میدهد حتی در غلظتهای بسیار پایین هم گلوکز منتقل میشود.
GLUT1
بیان این ناقل در بافتهای جنینی، گلبول قرمز و سد خونی-مغزی افراد بالغ و سلولهای توموری زیاد است. تعداد GLUT1 در غشا بستگی به غلظت گلوکز سلول دارد.
GLUT2
GLUT2 ورود گلوکز برای تشکیل گلیکوژن در کبد را تسهیل میکند و در پانکراس با انتقال دوطرفه گلوکز، سلولهای بتا را قادر به تعیین غلظت گلوکز خون میکند. برای جبران تمایل کم این ناقل به گلوکز، تعداد آن در غشا افزایش یافته است.
GLUT3
GLUT3 بیشتر در نورونها وجود دارد و تصور میشود بیشترین بخش جذب گلوکز در این سلولها را بر عهده دارد.
GLUT4
پس از مصرف غذا، سلولهای پانکراس انسولین ترشح میکنند. این هورمون، جذب گلوکز از خون توسط سلولهای ماهیچه اسکلتی و چربی را تسهیل میکند. اتصال انسولین به گیرنده آن، سبب ایجاد آبشار انتقال پیام در سلول و انتقال GLUT4 همراه وزیکولهای درونسلولی به غشای پلاسمایی سلولهای چربی و ماهیچهای میشود. همچنین، در زمان انقباض، ماهیچه اسکلتی نیاز به مصرف گلوکز بیشتر دارد و انتقال GLUT4 به غشا را تسهیل میکند.
SGLTs
SGLT (Sodium Glucose Linked Cotransporters) یک همانتقال است که سدیم و گلوکز را همزمان از غشا عبور میدهد. وابستگی این ناقلها به ATP غیرمستقیم و برای تنظیم غلظت یون سدیم سیتوپلاسمی است. ۶ ایزوفرم این ناقل در انسان شناخته شده است که درمورد SGLT1 و SGLT2 اطلاعات بیشتری داریم.
| نوع ناقل | اندام دارای ناقل |
ویژگی ناقل |
| SGLT1 | سلولهای اپیتلیوم روده |
عدم وابستگی به انسولین وابسته به ATP و |
| SGLT2 | توبول پروکسیمال نفرون |
عدم وابستگی به انسولین وابسته به ATP و |
انتقال فعال
انتقال فعال، به ناقل پروتئینی و انرژی ATP یا اختلاف غلظت الکتروشیمیایی برای انتقال مواد در دو طرف غشا نیاز دارد. در انتقال فعال اولیه، ۳ دسته پمپ ATPase نقش دارند که در ادامه آنها را بررسی میکنیم.
پمپهای ATPase
این پمپها مسئول انجام انتقال فعال اولیه هستند و انرژی حاصل از شکسته شدن پیوند فسفودیاستری در ATP را برای انتقال یونها بر خلاف شیب غلظت، استفاده میکنند. این پمپها براساس مکانسیم انتقال یونها در پستانداران به ۳ دسته تقسیم میشوند.
- پمپ P
- پمپ V
- پمپ F
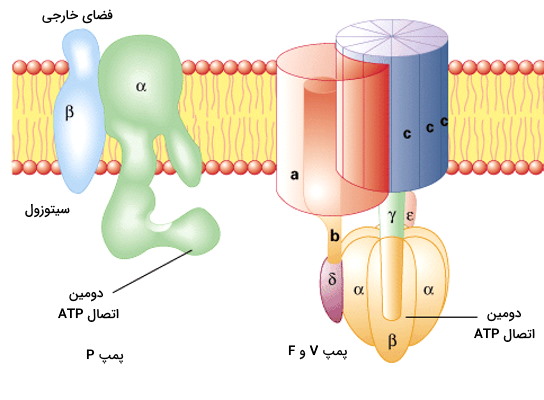
پمپ ATPase گروه P
برای بررسی عملکرد این دسته از ناقلها، پمپ «» و «» را در ادامه توضیح میدهیم.
پمپ
این پمپ برای بسیاری از فعالیتهای سلولی ازجمله تنظیم اسمزی بسیار مهم است. با مصرف یک مولکول ATP و با همانتقالی، ۳ یون سدیم را از سلول خارج و یک را به سلول وارد میکند. فسفوریله شدن باقیمانده آسپارات (Aspartate Residue) در این پمپ، باعث تغییر کنفورماسیون و انتقال یونها میشود. تغییر کنفورماسیون باعث میشود «تمایل» (Affinity) پمپ به داخل سلول و به خارج سلول، بیشتر باشد.
پمپهایی از این دست که در تنظیم ولتاژ غشا نقش دارند، «پمپهای الکتروژنیک» (Electrogenic Pump) نیز شناخته میشوند.
پمپ
پمپ از ۱۰ هلیکس عرض غشایی تشکیل شده است که به جز بخش اتصالی به ، همه آبگریز هستند. این پمپ به ازای مصرف هر ATP، ۲ را برخلاف شیب غلظت از سلول خارج میکند. تغییر کونفورماسیون در این پمپ نیز با فسفریله شدن باقیمانده آسپارات انجام میگیرد و باعث میشود تمایل پروتئین به در بخش سیتوپلاسمی بیشتر از خارج سلول باشد.
در سلولهای ماهیچهای، این پمپ بیشترین پروتئین موجود در شبکه اندوپلاسمی (۸۰ درصد پروتئینهای این غشا) را تشکیل میدهد و موجود در سیتوپلاسم را برای ذخیرهسازی وارد این اندامک میکند.
پمپ ATPase گروه V
پمپ ATPase گروه V در غشای واکوئلهای گیاهی، غشای لیزوزوم و اندوزوم جانوری، استئوکلاست و بعضی توبولهای کلیه قرار دارد. این پمپ و پمپهای گروه F از نظر ساختار یکسان هستند اما عملکرد متفاورتی دارند. پمپهای گروه V ناقل هستند و محیط اندامک را اسیدی میکنند. بسیاری از این پمپها، برای جبران اختلاف الکتریکی ناشی از انتقال ، با کانالهای جفت میشوند.
بلافاصله پس از ورود به لیزوزوم نیز وارد این اندامک میشود. در غیر این صورت، اشباع سیتوزول از ، سبب توقف فعالیت پمپ خواهد شد. همزمان پمپهای دیگر مثل ، را از لیزوزوم خارج و را وارد میکند تا پتانسیل عمل در غشا ایجاد نشود.
پمپ ATPase گروه F
این پمپ که ATP «سنتتاز» (ATP Synthase) و «ATP فسفوهیدرولاز» (ATP Phosphohydrolase) نیز نام دارد، کمپلکسی چند جزءی و در غشای داخلی میتوکندری قرار دارد. ATPase گروه F، از دو کانال و تشکیل شده است که با انتقال ، مولکول ATP تولید میکنند. این کانالها، موتورپروتئینهای چندزیرواحدی هستند که بهوسیله یک پروتئین حساس به اولیگومایسین بههم متصل شدهاند.

تفاوت انتشار تسهیل شده و انتقال فعال ثانویه
انتشار تسهیلشده مربوط به انتقال مواد از عرض غشا به کمک پروتئینهای غشایی است و اگر این انتقال برخلاف شیب غلظت و با صرف انرژی حاصل از گرادیان الکتروشیمیایی باشد، به آن انتقال فعال ثانویه میگویند.
نقش ناقلهای در تولید و ترشح اسید معده
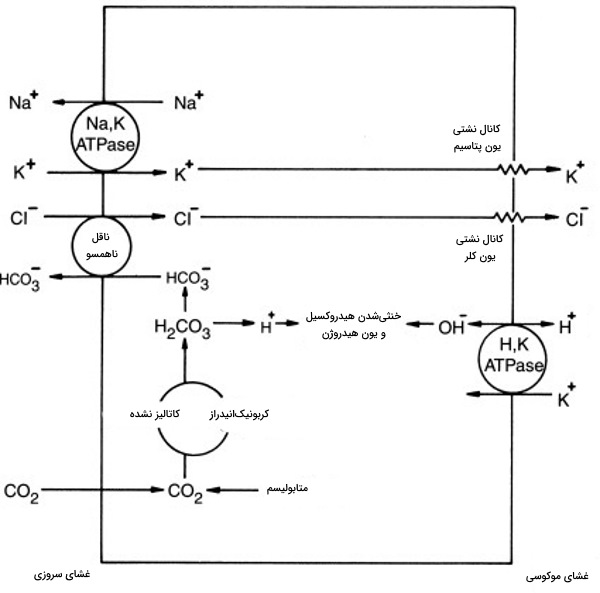
سنتز و ترشح اسید معده توسط «سلولهای پاریتال» (Parietal Cells) در دیواره معده انجام میشود و برای ورود املاح مورد نیاز در سنتز و ترشح اسید به حفره معده، به ۴ ناقل پروتئینی نیاز است.
پمپ پروتئینی عرض غشایی از نوع پمپهای P است. به بخش سیتوپلاسمی این ناقل آنزیمی متصل میشود و پس از تغییر کنفورماسیون پروتئین در اثر فسفوریله شدنِ باقیمانده آسپارتات، از سمت دیگر غشا خارج و پروتئین به متصل می شود. داروی «اُمپرازول» (Omeprazole) این پمپ را مهار میکند.
کانالهای نشتی و در غشای لومنی سلولهای پاریتال قرار دارند و به خروج و و حفظ تعادل غلظت سیتوپلاسمی این یونها کمک میکنند.
خروج ،pH اطراف سلولهای جداره معده را پایین میآورد. برای متعادل کردن pH محیط، ناقل آنیونی ،در سمت غشای پایه ساخته شده توسط آنزیم کربنیکانیدراز را از سلول خارج و را وارد سلول میکند. این انتقال بهصورت تبادل ناهمسوی یک یون در هربار فعالیت است.
آکوآپورین
«آکوآپورینها» (Aquaporins) پروتئینهای عرض غشایی هستند که به عنوان کانالهای انتقال آب در باکتریها، گیاهان و حیوانات عمل میکنند. در پستانداران تاکنون، در ایزوفرم این کانال شناخته شده است و هر کدام از این ایزوفرمها در اندام متفاوتی هستند. بهطور مثال، AQP0 در عدسی چشم و AQP1 در رگهای خونی و توبولهای کلیه قرار دارد. این کانالها از ۶ هلیکس عرض غشایی و ۲ هلیکس کوتاه خارج غشایی، تشکیل شده است. ساختار این کانال در غشا به صورت تترامر است. این کانالها و را نیز عبور میدهند.
آکوآپورین در نفرون
هفت نوع آکوآپورین در سلولهای کلیه بیان میشود و نقش مهمی در هومئوستازی بدن ایفا میکند. آکوآپورین۲ یکی از شناخته شدهترین این کانالهای آبی در نفرون است که به توضیح آن میپردازیم.
آکوآپورین ۲
آکوآپورین۲ (AQP2) کانال آبی است که در لولههای جمعکننده ادرار در نفرون قرار دارد و بهوسیله هورمونهای ADH و «وازوپرسین» (Vasopressin) کنترل میشود. در کمبود آب، بدن هورمون ADH ترشح میکنند. ADH آبشاری از واکنشهای درون سلولی را فعال میکند که در نهایت سبب ورود AQP2 به سطح غشای سلولها، در لوله جمعکننده ادرار میشود. این کانالها به بازجذب آب از نفرون و حفظ تعادل اسمزی بدن کمک میکنند. اختلال در ساختار یا عملکرد این کانال آبی، فرد را به دیابت بیمزه مبتلا میکند.
آکوآپورین غدد بزاقی
این کانالها در غدد بزاقی، آب را براساس اختلاف فشار اسمزی از غشا عبور میدهند. آکوآپورینهای بیان شده در این غدد انواع AQP1-AQP5 هستند. به عنوان نمونه، AQP5 در غشای «سلولهای آسینی» (Acinar cells) قرار دارد. پس از ترشح مایع غنی از توسط سلولهای آسینی، این آکوآپورین آب را به لومن آسینی منتقل میکند.
نفوذپذیری انتخابی در غشای هسته
بخشی از نفوذپذیری انتخابی غشای هسته مربوط به «منافذ هسته» (Nuclear Pores) میشود. منافذ هسته ساختاری شبیه به سبد دارند که از غشای داخلی و خارجی هسته گذر میکنند. این منافذ از مجموعه پروتئینی بزرگی - هر کدام ۳۰ پروتئین متفاوت - تشکیل شده است. منافذ هسته وظیفه انتقال آب و یونها و پروتئینهای دارای توالی هستهای را بر عهده دارد.
ساختار منافذ هسته
هر یک از منافذ هسته، مجموعهای از پروتئینها با ساختار متفاوت و ۸ وجهی است که تبادل مواد بین هسته و سیتوپلاسم از کانال استوانهای در مرکز آن انجام میشود. این ساختار از بخشهاس زیر تشکیل شده است.
- ۸ فیلامنت سیتوپلاسمی
- حلقه سیتوپلاسمی
- حلقه لومنی
- مجموعه میلهای
- حلقه هسته
- سبد هسته
- حلقه انتهایی
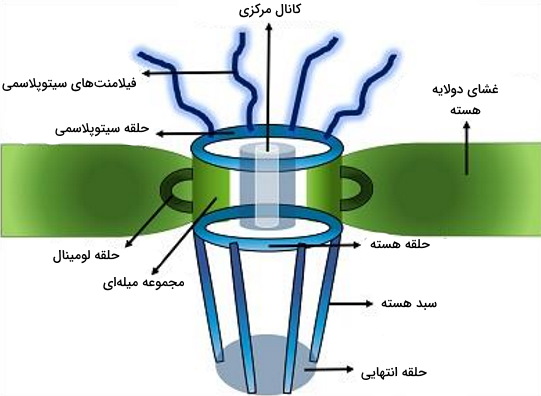
انتقال مواد از غشای هسته
ویژگی نفوذپذیری انتخابی هسته که توسط منافذ ایجاد میشود، ورود و خروج مولکولهای زیستی بالای ۵۰ کیلودالتون را با داشتن شناسههای منحصربهفردی از اسیدهای آمینه هسته محدود میکند. برای نمونه هیستون و RNA پلیمراز با «سیگنالهای مکانیابی هستهای» (Nuclear Localization Signals/NLS) نشانهگذاری شدهاند که جایگاه اتصال برای رسپتور هستهای «ایمپورتین» (Importin) است.
RNA و زیرواحدهای ریبوزوم با «سیگنالهای خروج از هسته» (Nuclear Export Signal/NES) نشانهگذاری شدهاند و این نشانه جایگاه اتصال به رسپتور هستهای «اکسپورتین» (Exportin) است. این نشانه توالی کوتاهی از اسیدآمینه لوسین است.
چرخه ورود مولکولها به هسته
رسپتور ایمپورتین دارای دو زیرواحد و است. مولکولی که میخواهد وارد هسته شود ازطریق NLS به به زیر واحد این رسپتور و همزمان کمپلکس RAN-GDP موجود در سیتوزول به زیر واحد متصل خواهد شد. سپس کمپلکس بهوجود آمده، با پروتئینهای منافذ هسته برهمکنش و وارد هسته میشود. مولکول ورودی، در هسته از کمپلکس RAN-GDP-Importin جدا و این کمپلکس به صورت RAN-GTP-Importin از منافذ هسته خارج میشود.
چرخه خروج مولکولها از هسته
شروع این چرخه با اتصال رسپتور اکسپورتین به توالی NES در مولکول خارجشونده از هسته همراه است. سپس با اتصال RAN-GTP به اکسپورتین، کل مجموعه با پروتينهای منفذ هسته برهمکنش میکند و از هسته خارج میشود. کمپلکس RAN-GAP موجود در سیتوزول، با گرفتن یک فسفات (P) و ایجاد تغییر در کنفورماسیون RAN-GTP مولکول مورد نظر را در سیتوزول آزاد میکند. این تبادلP، با کمک «فاکتور تبادل نوکلئوتید گوانین» (Guanine Nucleotide Exchange Factor) انجام میشود.
نفوذپذیری انتخابی غشا در سد خونی مغزی
سد خونی-مغزی غشای سلولهای اندوتلیال مویرگهای خونی است که برای در امان نگهداشتن سلولهای مغزی از هرگونه پاتوژن، نفوذپذیری انتخابی بسیار تخصصیافتهای دارد و تنها به تعداد کمی از مولکولها اجازه عبور از دولایه لیپیدی را میدهد. به همین دلیل نفوذپذیری بسیار کمی نسبت به مواد مختلف دارد و عملکرد متمایزی دارد. مولکولهای لیپیدی کوچک، تنها مولکولهای هستند که بهوسیله انتشار ساده از این سد عبور میکنند. ورود متابولیتهای کوچک و قطبی مثل گلوکز، آمینواسید، آنیونها و کاتیونهای آلی به سلولهای عصبی بهکمک این ناقلها انجام میگیرد . تخمینها نشان میدهند ۱۰ تا ۱۵ درصد پروتئینهای موجود در غشای مویرگهای عصبی، ناقلها هستند.
عوامل موثر بر نفوذپذیری غشا
نفوذپذیری یک غشا بهوسیله دما، نوع حلشوندههای موجود در دو طرف آن و هیدراته بودن سلول تحت تاثیر قرار میگیرد. افزایش دما باعث ناپایداری و افزایش سیالیت غشا و کاهش دما باعث سختتر شدن غشا میشود. دما، انرژی جنبشی غشا را بهوسیله تغییر در انرژی جنبشی فسفولیپیدها تغییر میدهد. حلشوندههای موجود، ناقلهای پروتئینی را تحت تاثیر قرار میدهند. ناقلهای پروتئینی عملکردهای مختلفی دارند که در این مقاله به آنها اشاره شد و اشباع شدن آنها از انتقال مواد بیشتر جلوگیری میکند. هر چه غلظت آب سلول کمتر باشد، نفوذپذیری غشا کمتر است.
جمعبندی
نفوذپذیری انتخابی ویژگی است که با غشای پلاسمایی سلول تعریف میشود. ساختار هیدروفوب حاصل از دولایه لیپیدی، از عبور مولکولهای بزرگ و ناقطبی در غشا جلوگیری میکند. به همین دلیل عبور مواد، نیاز به ساختارهای پروتئینی به نام ناقل دارد که با صرف انرژی یا بدون آن، مواد مورد نیاز برای متابولیسمهای سلولی را فراهم کند.









