



تنظیم عصبی چیست و عملکرد انواع سیستم عصبی چگونه است؟ – به زبان ساده
سیستم عصبی اصلیترین سیستم کنترلکننده، تنظیمکننده و ارتباط دهنده بدن است. مرکز تمام فعالیتهای ذهنی از جمله فکر، یادگیری و حافظه است. همراه با سیستم غدد درونریز، سیستم عصبی مسئول تنظیم و حفظ هومئوستاز است. در این مقاله به طور کامل به بررسی نحوه تنظیم عصبی انسان و عملکردهای سیستمهای عصبی و همچنین انواع رفلکسهای مختلف در بدن انسان میپردازیم.


تنظیم عصبی چیست؟
سیستم عصبی از طریق گیرندههای خود، ما را با محیط خود، چه بیرونی و چه داخلی، در تماس نگه میدارد. مانند سایر سیستمهای بدن، سیستم عصبی از اندامهایی تشکیل شده است که عمدتاً مغز، نخاع، اعصاب و گانگلیونها هستند. اینها به نوبه خود از بافتهای مختلفی از جمله عصب، خون و بافت همبند تشکیل شدهاند. اینها با هم فعالیتهای پیچیده تنظیم عصبی را انجام میدهند. فعالیتهای مختلف تنظیم عصبی را میتوان با هم به عنوان سه عملکرد کلی و قابل همپوشانی حسی، حرکتی و یکپارچه دستهبندی کرد.
میلیونها گیرنده حسی، تغییراتی به نام محرک را که در داخل و خارج بدن رخ میدهند، تشخیص میدهند. آنها مواردی مانند دما، نور و صدا را از محیط بیرونی کنترل میکنند. در داخل بدن گیرندههای محیط داخلی مواردی مثل تغییرات فشار، pH، غلظت دیاکسید کربن و سطوح الکترولیتهای مختلف را تشخیص میدهند. همه این اطلاعات جمعآوری شده، ورودی حسی نامیده شده و به تنظیم عصبی منجر میشود.
تعریف سیستم عصبی
اصلیترین سیستم کنترلکننده، تنظیمکننده و ارتباطدهنده بدن بوده و مرکز تمام فعالیتهای ذهنی از جمله فکر، یادگیری و حافظه است. همراه با سیستم غدد درونریز، سیستم عصبی مسئول تنظیم عصبی و حفظ هومئوستاز است. این سیستم از طریق گیرندههای خود ما را با محیط خود، چه بیرونی و چه داخلی، در تماس نگه میدارد. مدارهایی که تابع عملکردهای مشابه هستند در سیستمهای عصبی گروهبندی میشوند که اهداف رفتاری گستردهتری را انجام میدهند. کلیترین تعریف کاربردی، سیستمهای عصبی را به سیستمهای حسی مانند بینایی یا شنوایی تقسیم میکند که اطلاعات را از محیط دریافت و پردازش میکنند، و همچنین سیستمهای حرکتی که به ارگانیسم اجازه میدهد با ایجاد حرکات به چنین اطلاعاتی پاسخ دهد.
ورودی حسی به سیگنالهای الکتریکی به نام تکانههای عصبی تبدیل میشود که به مغز منتقل میشود. در آنجا سیگنالها برای ایجاد احساسات، ایجاد افکار یا افزودن به حافظه، گرد هم میآیند. هر لحظه بر اساس ورودی حسی تصمیمگیری میشود، این حالت بیانگر یکپارچگی است. بر اساس ورودی حسی و یکپارچگی، سیستم عصبی با ارسال سیگنال به عضلات، باعث انقباض آنها یا به غدد، که باعث تولید ترشحات میشود، پاسخ میدهد. ماهیچهها و غدد را افکتور مینامند زیرا در پاسخ به دستورات تنظیم عصبی اثری ایجاد میکنند. این خروجی، «موتور» یا «عملکرد حرکتی» نام دارد.

سیستم عصبی از چه قسمت هایی تشکیل شده است؟
دستگاه عصبی به طور کلی به دو بخش تقسیم میشود: سیستم عصبی مرکزی (CNS) و سیستم عصبی محیطی (PNS). هر بخش دارای ویژگیهای ساختاری و عملکردی است که آن را از سایرین متمایز میکند. اگرچه هر بخش فرعی این سیستم یک سیستم تنظیم عصبی نیز نامیده میشود، اما همه این سیستمهای کوچکتر به یک سیستم عصبی منفرد و بسیار یکپارچه تعلق دارند. در ادامه به هر کدام از بخشهای این سیستم و اجزای آنها میپردازیم.
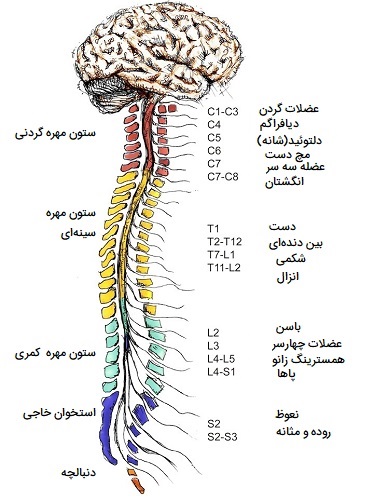
سیستم عصبی مرکزی چیست؟
CNS یا Central Nervous System شامل مغز و نخاع است که در حفره پشتی بدن قرار دارند. مغز توسط جمجمه احاطه شده است و نخاع توسط مهرهها محافظت میشود. مغز با طناب نخاعی در فورامن مگنوم (حفره بزرگ جمجمه) پیوسته است. علاوه بر استخوان، CNS توسط غشای بافت همبند به نام مننژ و مایع مغزی - نخاعی احاطه شده است. در ادامه اجزای مختلف سیستم عصبی مرکزی را بررسی میکنیم.
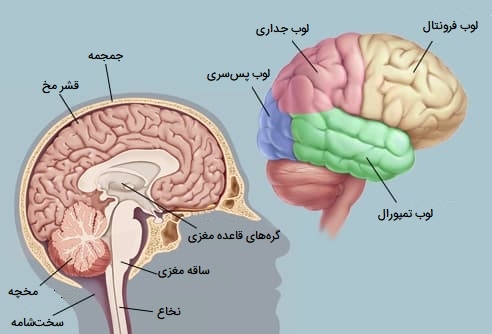
مغز
مغز به قشر مخ، دیانسفالون، ساقه مغز و مخچه تقسیم میشود. در ادامه هر کدام را بیشتر بررسی کردهایم.
- «قشر مخ» (Cerebrum). بزرگترین و واضحترین بخش مغز مخ است که توسط یک شکاف طولی عمیق به دو نیمکره مغزی تقسیم میشود. دو نیمکره دو موجود مجزا هستند اما توسط یک نوار قوسدار از الیاف سفید به نام جسم پینهای که مسیر ارتباطی بین دو نیمه را فراهم میکند به هم متصل شده اند. هر نیمکره مغزی به پنج لوب تقسیم میشود که چهار تای آنها نامی مشابه استخوان روی آنها دارند. لوب فرونتال (پیشانی)، لوب جداری (پریتال)، لوب اکسیپیتال (پسسری) و لوب تمپورال. لوب پنجم، اینسولا یا جزیره ریل، در اعماق شیار جانبی قرار دارد.
- «دیانسفالون» (Diencephalon). دیانسفالون ناحیهای از لوله عصبی جنینی مهرهداران است که ساختارهای قدامی جلوی مغز از جمله تالاموس، هیپوتالاموس و غیره را ایجاد میکند. دیانسفالونها در مرکز قرار دارند و تقریباً توسط نیمکرههای مغزی احاطه شدهاند. تالاموس، حدود 80 درصد از دیانسفالونها را شامل شده و از دو توده بیضی شکل از ماده خاکستری تشکیل شده است که به عنوان ایستگاههایی برای تکانههای حسی، به جز حس بویایی انسان که به قشر مغز میرود، عمل میکنند. هیپوتالاموس ناحیه کوچکی در زیر تالاموس است که نقش کلیدی در حفظ هومئوستاز دارد زیرا بسیاری از فعالیتهای احشایی را تنظیم میکند. این غده کوچک با شروع بلوغ و چرخههای ریتمیک در بدن درگیر است و مانند یک ساعت بیولوژیکی عمل میکند.
- «ساقه مغز» (Brain Stem). ساقه مغز ناحیه بین دیانسفالون و نخاع است. از سه قسمت تشکیل شده است: «مغز میانی» (midbrain)، «پل مغزی» (pons) و «بصل النخاع» (medulla oblongata). مغز میانی بالاترین بخش ساقه مغز است. پل مغزی قسمت میانی برآمده ساقه مغز است. این ناحیه عمدتاً از رشتههای عصبی تشکیل شده است که مجاری رسانایی را بین مراکز بالاتر مغز و نخاع تشکیل میدهند. بصل النخاع یا مدولا، در قسمت پایینی از پل مغزی امتداد مییابد. با طناب نخاعی در فورامن مگنوم پیوسته است. تمام رشتههای عصبی صعودی (حسی) و نزولی (حرکتی) که مغز و نخاع را به هم متصل میکنند از بصلالنخاع عبور میکنند.
- «مخچه» (Cerebellum). مخچه، دومین بخش بزرگ مغز، در زیر لوبهای پس سری مخ قرار دارد. سه دسته جفتی از رشتههای عصبی میلیندار، به نام دمگلهای مخچه، مسیرهای ارتباطی بین مخچه و سایر قسمتهای سیستم عصبی مرکزی را تشکیل میدهند. مخچه مرکز مهمی برای هماهنگی حرکات و کنترل تعادل و وضعیت بدن است.
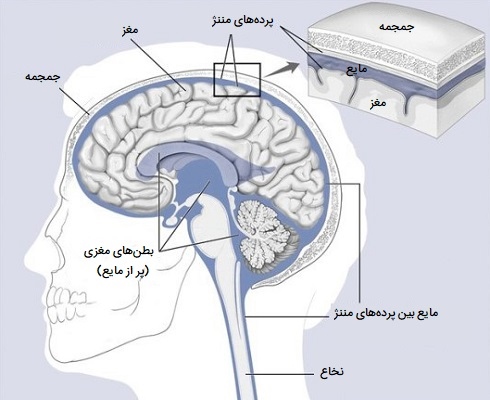
پرده مننژ چیست؟
سه لایه مننژ در اطراف مغز و نخاع وجود دارد. لایه بیرونی، سختشامه، بافت همبند فیبری سفید و سفت است. لایه میانی مننژ عنکبوتیه است که از نظر ظاهری شبیه تار عنکبوت است، یک لایه نازک با رشتههای نخ مانند متعدد است که آن را به درونیترین لایه متصل میکند. فضای زیر عنکبوتیه، با مایع مغزی - نخاعی پر شده و حاوی رگهای خونی است. «نرمشامه» (The pia mater) داخلیترین لایه مننژ است. این غشای نازک و ظریف به شدت به سطح مغز و نخاع متصل است و بدون آسیب رساندن به سطح قابل جدا شدن نیست.
مننژیومها تومورهای بافت عصبی پوشاننده مغز و نخاع هستند. اگرچه مننژیومها معمولاً به احتمال زیاد گسترش نمییابند، پزشکان اغلب برای درمان علائمی که ممکن است با اعمال فشار تومور به مغز ایجاد شوند، با آنها طوری رفتار میکنند که گویی بدخیم هستند.
بطن ها و مایع مغزی نخاعی
مجموعهای از حفرههای به هم پیوسته و پر از مایع در مغز یافت میشود. این حفرهها بطنهای مغز هستند و مایع آن مایع مغزی - نخاعی (CSF) است. مغز به شدت به تامین مداوم گلوکز و اکسیژن از طریق خون وابسته است به این دلیل که گلیکوژن ذخیره شده کمی دارد. تبادل مواد بین خون و مایع خارج سلولی در CNS از طریق گروه پیچیدهای از مکانیسمهای سد خونی - مغزی بسیار محدود شده است. مایع مغزی - نخاعی و مایع خارج سلولی در مغز در تعادل انتشاری با یکدیگر هستند اما تفاوت را با خون حفظ میکنند.
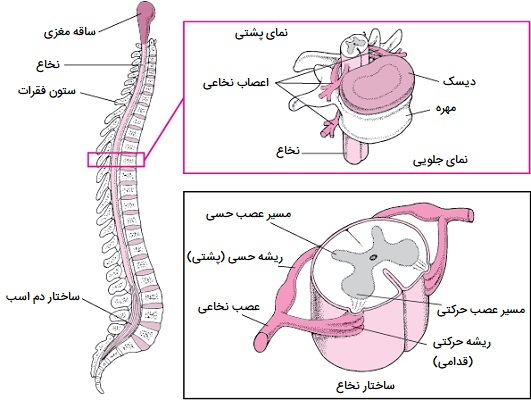
نخاع
طناب نخاعی از فورامن مگنوم در پایه جمجمه تا سطح اولین مهره کمری امتداد دارد. طناب با بصل النخاع در فورامن مگنوم پیوسته است. مانند مغز، نخاع نیز توسط استخوان، مننژها و مایع مغزی نخاعی احاطه شده است. 31 جفت اعصاب نخاعی با 4 سطح خروجی مشخص میشوند که شامل اعصاب گردنی (8)، سینهای (12)، کمری (5) و خاجی (5) هستند. فیبرهای آوران از سیستم محیطی وارد قسمت پشتی بند ناف از طریق ریشههای پشتی شده و گرههای ریشه پشتی را تشکیل میدهند. فیبرهای وابران از طریق ریشههای شکمی، بند ناف را در سمت شکمی ترک میکنند.
ریشههای پشتی و شکمی از یک سطح با هم ترکیب میشوند و یک عصب نخاعی در خارج از طناب (یکی در هر طرف) را تشکیل میدهند. در انتهای دیستال طناب، بسیاری از اعصاب نخاعی فراتر از مخروط مدولاریس گسترش یافته و مجموعهای شبیه دم اسب را تشکیل میدهند. این قسمت دم اسب یا cauda equina است و در مقطع عرضی، طناب نخاعی بیضی شکل به نظر میرسد. نخاع دو وظیفه اصلی دارد:
- به عنوان یک مسیر هدایت برای تکانههایی که به مغز میروند و از آن خارج میشوند. تکانههای حسی در مسیرهای صعودی ستون فقرات به مغز میروند. تکانههای حرکتی در مسیرهای نزولی حرکت میکنند.
- همچنین نخاع به عنوان یک مرکز رفلکس خدمت میکند. قوس رفلکس واحد عملکردی تنظیم عصبی است. رفلکسها پاسخ به محرکهایی هستند که نیازی به تفکر آگاهانه ندارند و در نتیجه سریعتر از واکنشهایی که نیاز به فرآیندهای فکری دارند رخ میدهند. به عنوان مثال، با رفلکس خروج، عمل رفلکس قبل از اینکه از درد آگاه شوید، قسمت آسیب دیده را خارج میکند. بسیاری از رفلکسها در نخاع بدون رفتن به مراکز بالاتر مغز واسطه میشوند.
سیستم عصبی محیطی
سیستم عصبی محیطی شامل اعصابی است که از مغز و نخاع منشعب میشوند. این اعصاب شبکه ارتباطی بین CNS و اعضای بدن را تشکیل میدهند. سیستم عصبی محیطی خود به سیستم عصبی سوماتیک و سیستم عصبی خودمختار تقسیم میشود.
سیستم عصبی پیکری یا سوماتیک
«سیستم عصبی سوماتیک» (Somatic Nervous System) متشکل از اعصابی است که به سمت پوست و ماهیچهها میروند و در تنظیم عصبی فعالیتهای آگاهانه شرکت دارند. این سیستم عضلات اسکلتی را عصبدهی میکند و از آکسونهای میلیندار بدون هیچ سیناپسی تشکیل شده است. فعالیت این نورونها منجر به تحریک (انقباض) عضلات اسکلتی میشود و به همین دلیل به آنها نورونهای حرکتی میگویند، این نوع نورونها هرگز بازدارنده نیستند.
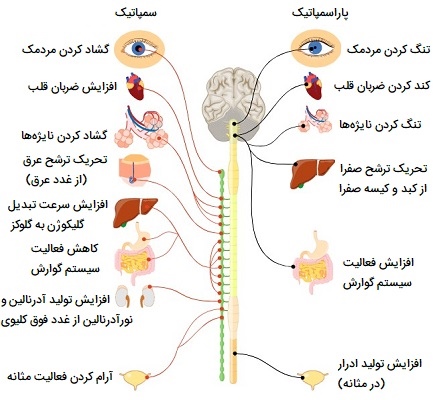
سیستم عصبی خودمختار
«سیستم عصبی خودمختار» (Autonomic Nervous System) یک سیستم وابران احشایی است، به این معنی که تکانههای حرکتی را به اندامهای احشایی میفرستد. این سیستم به طور خودکار و مداوم، بدون تلاش آگاهانه، برای عصبدهی به عضلات صاف، ماهیچههای قلب و غدد عمل میکند. اعمال این سیستم مربوط به ضربان قلب، ضربان تنفس، فشار خون، دمای بدن و سایر فعالیتهای احشایی است که برای حفظ هومئوستاز با هم کار میکنند.
سیستم عصبی خودمختار شامل اعصابی است که سیستم عصبی مرکزی را به اندامهای احشایی مانند قلب، معده و رودهها متصل میکند و واسطه فعالیتهای تنظیم عصبی ناخودآگاه است. زنجیرههای موازی که هر کدام دارای دو نورون هستند، سیستم عصبی مرکزی و سلولهای عامل را به هم متصل میکنند. سیناپس بین این دو نورون را گانگلیون خودمختار، رشتههای عصبی بین CNS و گانگلیون را فیبرهای پیشگانگلیونی و رشتههای بینگانگلیون و سلولهای عامل را فیبرهای پسگانگلیونی میگویند.
این نوع سیستم عصبی به دو نوع تنظیم عصبی سمپاتیک (جنگ یا فرار) و پاراسمپاتیک (استراحت و آرامش) تقسیم میشود. گرههای سمپاتیک نزدیک به نخاع و گرههای پاراسمپاتیک نزدیک به اندامها قرار دارند. بسیاری از اندامهای احشایی با رشتههای عصبی از هر دو بخش تامین میشوند. در این صورت در هنگام تنظیم عصبی یکی تحریک و دیگری مهار را موجب میشود. این رابطه عملکردی متضاد به عنوان تعادلی برای کمک به حفظ هومئوستاز عمل میکند. در ادامه به تنظیم عصبی سمپاتیک و پاراسمپاتیک بیشتر میپردازیم.
- سیستم سمپاتیک طوری تنظیم شده است که به عنوان یک واحد عمل کند در حالی که سیستم پاراسمپاتیک به گونهای مرتب شده است که قسمتها بتوانند به طور مستقل عمل کنند.
- سیستم سمپاتیک در پاسخ به استرس نقش دارد.
- بسیاری از اندامها و غدد یک عصب دوگانه از رشتههای سمپاتیک و پاراسمپاتیک دریافت میکنند.
- این دو سیستم عموماً اثرات متضادی دارند و با هم برای تنظیم عصبی یک پاسخ کار میکنند. اغلب پاسخهای خودمختار معمولاً بدون کنترل آگاهانه رخ میدهند.
عصب چیست؟
یک عصب شامل دستههایی از رشتههای عصبی، آکسون یا دندریت است که توسط بافت همبند احاطه شدهاند. اعصاب حسی فقط شامل رشتههای آوران، دندریتهای بلند نورونهای حسی هستند. اعصاب حرکتی فقط دارای فیبرهای وابران هستند، یعنی آکسونهای بلند نورونهای حرکتی. اعصاب مختلط حاوی هر دو نوع فیبر هستند. یک غلاف بافت همبند به نام «اپی نوریوم» (The epineurium) هر عصب را احاطه کرده است.
هر دسته از رشتههای عصبی فاسیکلوس نامیده میشود و توسط لایهای از بافت همبند به نام «پری نوریوم» (The perineurium) احاطه شده است. در داخل فاسیکولوس، هر فیبر عصبی منفرد، با میلین و نوریلم خود، توسط بافت همبند به نام «اندونوریوم» (The endoneurium) احاطه شده است. یک عصب همچنین ممکن است دارای رگهای خونی باشد که در پوششهای بافت همبند خود محصور شدهاند.
اعصاب جمجمه ای
12 جفت اعصاب جمجمهای از سطح تحتانی مغز خارج میشوند. همه این اعصاب، به جز عصب واگ، از سوراخهای جمجمه عبور میکنند تا ساختارهای سر، گردن و ناحیه صورت را عصبدهی کنند. اعصاب جمجمهای هم با نام و هم با اعداد رومی بر اساس ترتیب ظاهر شدن آنها در سطح تحتانی مغز مشخص میشوند. بیشتر اعصاب دارای اجزای حسی و حرکتی هستند. سه تا از اعصاب با حس بویایی، بینایی، شنوایی و تعادل خاص مرتبط هستند و فقط دارای رشتههای حسی هستند. پنج عصب دیگر عمدتاً دارای عملکرد حرکتی هستند، اما دارای فیبرهای حسی برای حس عمقی هستند. چهار عصب باقی مانده از مقادیر قابل توجهی فیبرهای حسی و حرکتی تشکیل شده است.
نورومهای آکوستیک تودههای فیبری خوش خیمی هستند که از عصب تعادلی به وجود میآیند که عصب هشتم جمجمه یا عصب دهلیزی نیز نامیده میشود. این تومورها غیر بدخیم هستند، به این معنی که به سایر قسمتهای بدن گسترش نمییابند و متاستاز نمیدهند. محل این تومورها در عمق جمجمه و در مجاورت مراکز حیاتی مغز در ساقه مغز است. همانطور که تومورها بزرگ میشوند، ساختارهای اطراف را درگیر میکنند که با عملکردهای حیاتی ارتباط دارند.
در اکثر موارد، این تومورها در طی چند سال به کندی رشد میکنند. در موارد دیگر، سرعت رشد سریعتر است و بیماران علائم را با سرعت بیشتری بروز میدهند. معمولاً علائم خفیف هستند و بسیاری از بیماران تا مدتی پس از ایجاد تومور تشخیص داده نمیشوند. بسیاری از بیماران همچنین در طی چندین سال رشد توموری را نشان نمیدهند که اسکنهای سالیانه MRI برای آنها انجام میشود.
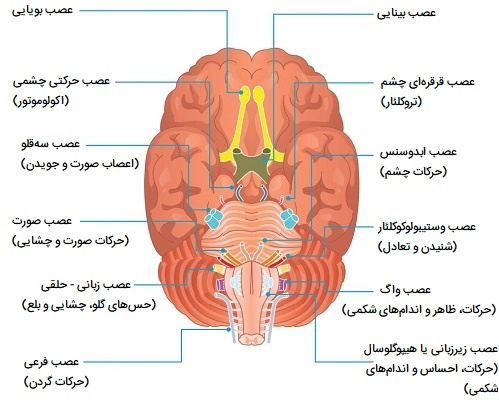
اعصاب نخاعی
سی و یک جفت اعصاب نخاعی به صورت جانبی از نخاع خارج میشوند. هر جفت عصب مربوط به قسمتی از بند ناف است و بر این اساس نامگذاری میشوند. این بدان معناست که 8 عصب گردنی، 12 عصب سینهای، 5 عصب کمری، 5 عصب ساکرال و 1 عصب دنبالچهای وجود دارد. هر عصب نخاعی توسط یک ریشه پشتی و یک ریشه شکمی به نخاع متصل میشود. اجسام سلولی نورونهای حسی در گانگلیون ریشه پشتی قرار دارند، اما اجسام سلولی نورون حرکتی در ماده خاکستری قرار دارند. این دو ریشه درست قبل از خروج عصب از ستون مهرهها به هم میپیوندند و عصب نخاعی را تشکیل میدهند. از آنجایی که همه اعصاب نخاعی دارای اجزای حسی و حرکتی هستند، همه آنها اعصاب مختلط هستند.
بافت عصبی چیست؟
اگرچه سیستم عصبی بسیار پیچیده است، اما تنها دو نوع سلول اصلی در بافت عصبی وجود دارند. سلول عصبی واقعی نورون است. این سلول رسانا است که تکانهها و واحد ساختاری سیستم عصبی را منتقل میکند. نوع دیگر سلول نوروگلیا یا سلول گلیال است. کلمه نوروگلیا به معنای چسب اعصاب است. این سلولها نارسانا هستند و یک سیستم پشتیبانی برای نورونها فراهم میکنند. آنها نوع خاصی از بافت همبند برای سیستم عصبی هستند. در ادامه انواع سلولهای بافت عصبی را بررسی میکنیم.
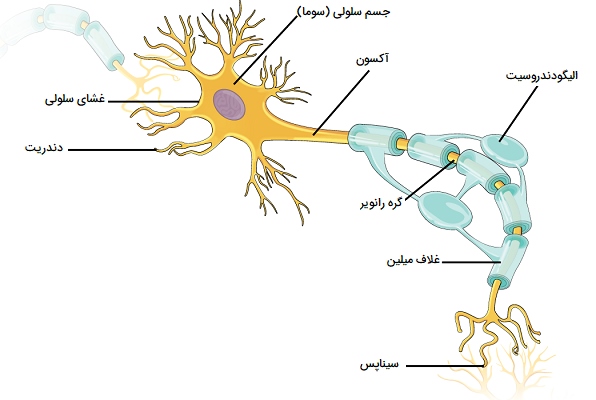
نورون چیست؟
نورونها یا سلولهای عصبی، وظایف سیستم عصبی را با هدایت تکانههای عصبی انجام میدهند. آنها بسیار تخصصی و آمیتوتیک هستند. آمیتوتیک بدان معنی است که اگر یک نورون از بین برود، نمیتوان آن را جایگزین کرد زیرا نورونها از طریق میتوز تقسیم نمیشوند. تصویر زیر ساختار یک نورون معمولی را نشان میدهد. هر نورون دارای سه بخش اصلی است: جسم سلولی (سوما)، یک یا چند دندریت و یک آکسون. بخشهای مختلف یک نورون و قسمتهای ساختاری آن را در ادامه بررسی کردهایم.
جسم سلولی
از بسیاری جهات، جسم سلولی مشابه انواع دیگر سلولها است. دارای حداقل یک هسته و حاوی بسیاری از اندامکهای سیتوپلاسمی معمولی بوده با این حال، فاقد سانتریول است. از آنجایی که سانتریولها در تقسیم سلولی عمل میکنند، این واقعیت که نورونها فاقد این اندامکها هستند، با ماهیت آمیتوزی (ناتوانی در تقسیم میتوز) سلول سازگار است.
دندریت ها
دندریتها و آکسونها امتداد سیتوپلاسمی یا فرآیندهایی هستند که از جسم سلولی بیرون میآیند. گاهی اوقات به آنها الیاف نیز گفته میشود. دندریتها معمولاً، اما نه همیشه، کوتاه و منشعب هستند که باعث افزایش سطح مقطع آنها برای دریافت سیگنال از نورونهای دیگر میشود. تعداد دندریتهای روی یک نورون متفاوت است. آنها فرآیندهای آوران نامیده میشوند زیرا تکانهها را به جسم سلولی عصبی منتقل میکنند. تنها یک آکسون وجود دارد که از هر جسم سلولی بیرون میزند. آکسون معمولاً دراز است و چون تکانهها را از جسم سلولی دور میکند، فرآیند وابران نامیده میشود.
آکسون
یک آکسون ممکن است شاخههای نادری به نام ضمائم آکسونی داشته باشد. آکسونها و ضمائم آکسون به بسیاری از شاخههای کوتاه یا تلودندری ختم میشوند. بخش انتهایی تلودندری کمی بزرگ شده و پیازهای سیناپسی را تشکیل میدهد. بسیاری از آکسونها توسط یک ماده چربی تقسیم شده، سفید رنگ به نام میلین یا غلاف میلین احاطه شده اند. فیبرهای میلیندار ماده سفید در سیستم عصبی مرکزی را تشکیل میدهند، در حالی که اجسام سلولی و الیاف غیر میلیندار ماده خاکستری را میسازند. نواحی بدون میلین بین بخشهای میلین را «گرههای رانویر» (The nodes of Ranvier) مینامند.
در سیستم عصبی محیطی، میلین توسط سلولهای شوان تولید میشود. سیتوپلاسم، هسته و غشای سلولی بیرونی سلول شوان پوشش محکمی در اطراف میلین و اطراف خود آکسون در گرههای رانویر تشکیل میدهند. این پوشش همان نوریلما است که نقش مهمی در بازسازی رشتههای عصبی دارد. در سیستم عصبی مرکزی، الیگودندروسیتها میلین تولید میکنند، اما نوریلم وجود ندارد، به همین دلیل است که فیبرهای درون CNS بازسازی نمیشوند.
از نظر عملکردی، نورونها بر اساس جهتی که در آن تکانهها را نسبت به سیستم عصبی مرکزی منتقل میکنند، بهعنوان آوران، وابران یا بیننورونی (نرونهای رابط) طبقهبندی میشوند. نورونهای آوران یا حسی، تکانههایی را از گیرندههای حسی محیطی به سیستم عصبی مرکزی حمل میکنند. آنها معمولاً دندریتهای بلند و آکسونهای نسبتاً کوتاه دارند. نورونهای وابران یا موتور، تکانههایی را از CNS به اندامهای مؤثر مانند ماهیچهها و غدد منتقل میکنند. نورونهای وابران معمولاً دندریتهای کوتاه و آکسونهای بلند دارند.
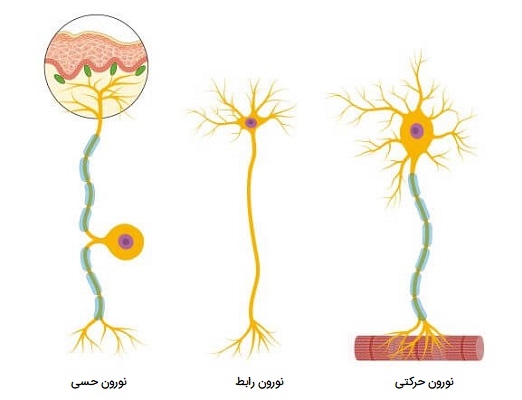
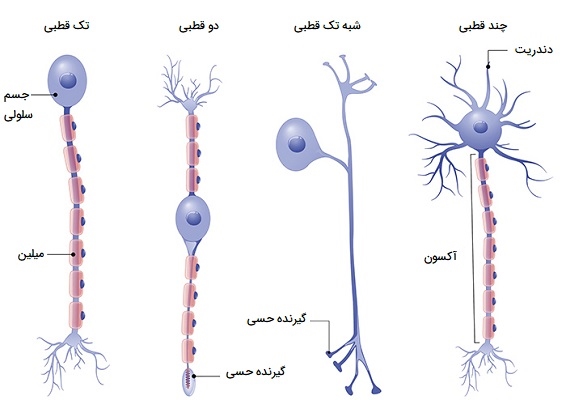
انواع نورون
اگرچه میلیاردها نورون و تغییرات گسترده وجود دارد، نورونها را میتوان بسته به عملکرد تنظیم عصبی آنها به سه گروه اصلی طبقهبندی کرد. نورونهای حسی (دندریتهای بلند و آکسونهای کوتاه)، نورونهای حرکتی (دندریتهای کوتاه و آکسونهای بلند) و نورونهای رابط (دندریتهای کوتاه و آکسونهای کوتاه یا بلند). در ادامه هر کدام از این نورونها را بیشتر بررسی میکنیم.
نورون حسی
نورونهای حسی (گاهی اوقات به عنوان نورونهای آوران شناخته میشوند) سلولهای عصبی هستند که تکانههای عصبی را از گیرندههای حسی به سمت سیستم عصبی مرکزی و مغز حمل میکنند. هنگامی که این تکانههای عصبی به مغز میرسند، به احساس مانند بینایی، شنوایی، چشایی و حس لامسه تبدیل میشوند. این اطلاعات حسی میتواند فیزیکی باشد، از طریق صدا، گرما، لمس و نور، یا میتواند از طریق طعم یا بو شیمیایی باشد. هنگامی که این اتفاق میافتد، نورونهای حسی سیگنالهایی را در مورد اطلاعاتی که دریافت کردهاند به سیستم عصبی مرکزی ارسال میکنند. اکثر نورونهای حسی شبه تکقطبی هستند. این بدان معنی است که آنها یک آکسون دارند که به دو شاخه تقسیم میشود.
نورون حرکتی
نورونهای حرکتی (که به عنوان نورونهای وابران نیز شناخته میشوند) سلولهای عصبی هستند که مسئول انتقال سیگنالها از سیستم عصبی مرکزی به سمت عضلات برای ایجاد حرکت هستند. آنها انتقال دهندههای عصبی را آزاد میکنند تا پاسخهایی را که منجر به حرکت ماهیچهها میشود، تحریک کنند. نورونهای حرکتی در ساقه مغز یا نخاع (بخشهایی از سیستم عصبی مرکزی) قرار دارند و به عضلات، غدد و اندامهای سراسر بدن متصل میشوند. نورونهای حرکتی به عنوان چند قطبی مشخص میشوند. این بدان معنی است که آنها یک آکسون و چندین دندریت دارند که از جسم سلولی بیرون میزند.
این نوع نورونها سیگنالهایی را از نخاع و ساقه مغز به عضلات اسکلتی و صاف منتقل میکنند تا به طور مستقیم یا غیرمستقیم حرکات ماهیچهها را کنترل کنند. به عنوان مثال، پس از لمس یک سطح داغ با دست، پیام از نورونهای حسی دریافت شده است. سپس نورونهای حرکتی باعث دور شدن دست از سطح داغ میشوند. دو نوع نورون حرکتی وجود دارد که شامل موارد زیر هستند:
- نورونهای حرکتی تحتانی: اینها نورونهایی هستند که از نخاع به انواع ماهیچههای بدن حرکت میکنند.
- نورونهای حرکتی فوقانی: این گروه از نورونهای حرکتی آنهایی هستند که بین مغز و نخاع حرکت میکنند.
نورون رابط
یک نورون رابط (همچنین به عنوان یک نورون داخلی شناخته میشود) به نورونهای حسی و حرکتی اجازه میدهد تا با یکدیگر ارتباط برقرار کنند. نورونهای رابط نورونهای مختلفی را در مغز و نخاع به هم متصل میکنند و به دلیل آکسون کوتاهشان به راحتی قابل تشخیص هستند. مانند نورونهای حرکتی، نورونهای بین عصبی چند قطبی هستند. این بدان معناست که آنها یک آکسون و چندین دندریت دارند. علاوه بر این که به عنوان یک اتصال بین نورونها عمل میکنند، نورونهای بین عصبی میتوانند از طریق تشکیل مدارهایی با پیچیدگیهای متفاوت با یکدیگر ارتباط برقرار کنند. ارتباط بین نورونها به مغز کمک میکند تا عملکردهای پیچیدهای مانند یادگیری و تصمیمگیری را انجام دهد، و همچنین نقشی حیاتی در رفلکسها و نوروژنز (بازسازی نورونهای جدید) بازی میکند.
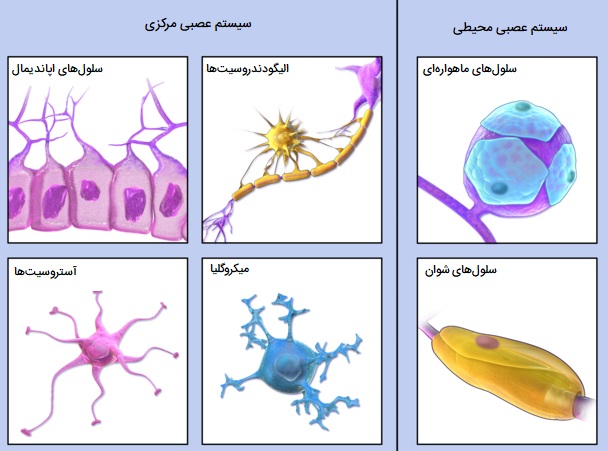
انواع سلول پشتیبان بافت عصبی
نوروگلیا که سلول گلیال یا گلیا نیز نامیده میشود، هر یک از چندین نوع سلولی است که عمدتاً برای حمایت از نورونها عمل میکند. اصطلاح نوروگلیا به معنای چسب عصبی است. در سال 1907، امیلیو لوگارو، زیستشناس ایتالیایی پیشنهاد کرد که سلولهای نوروگلیال مواد را با مایع خارج سلولی مبادله میکنند و از این طریق بر محیط عصبی کنترل دارند. از آن زمان نشان داده شده است که گلوکز، اسیدهای آمینه و یونها که همگی بر عملکرد نورون تأثیر میگذارند بین فضای خارج سلولی و سلولهای نوروگلیال مبادله میشوند. به عنوان مثال، پس از سطوح بالای فعالیت عصبی، سلولهای نوروگلیال میتوانند یونهای پتاسیم را جذب کرده و به صورت فضایی بافر ایجاد و در نتیجه عملکرد نورون طبیعی را حفظ کنند. نوروگلیا از تعداد نورونهای سیستم عصبی حداقل 10 برابر بیشتر است.
نوروگلیا در سیستم عصبی بیمهرگان و همچنین مهرهداران وجود دارد و با کمبود آکسون و وجود تنها یک نوع زائده از نورونها قابل تشخیص است. علاوه بر این، سیناپسها را تشکیل نمیدهند و توانایی تقسیم شدن را در طول عمر خود حفظ میکنند. در حالی که نورونها و نوروگلیا در نزدیکی یکدیگر قرار دارند، هیچ اتصال مستقیمی، مانند اتصالات شکافدار، بین این دو نوع وجود ندارد. «اتصالات شکافدار» (Gap junctions) بین سلولهای نوروگلیال وجود دارد. سلولهای نوروگلیا تکانههای عصبی را هدایت نمیکنند، اما در عوض از نورونها حمایت، تغذیه و محافظت میکنند. این سلولها برخلاف نورونها قادر به میتوز هستند.
جدا از تکنیکهای بافتشناسی و میکروسکوپی الکترونی مرسوم، تکنیکهای ایمونولوژیک برای شناسایی انواع مختلف سلولهای نوروگلیال استفاده میشوند. با رنگآمیزی سلولها با آنتیبادیهایی که به اجزای پروتئینی خاص نوروگلیاهای مختلف متصل میشوند، چند نوع نوروگلیا در سیستم عصبی مرکزی یافت میشود که شامل میکروگلیاها و ماکروگلیاها (که خود به انواع سلولهای دیگری تقسیم میشوند) هستند، در ادامه هر کدام را بررسی کردهایم.
میکروگلیا چیست؟
«میکروگلیا» (Microglia) سلولهای ایمنی مغز هستند که از آن در برابر آسیب و بیماری محافظت میکنند. میکروگلیا تشخیص میدهد که چه زمانی مشکلی پیش آمده است و پاسخی را آغاز کرده که عامل سمی را حذف یا سلولهای مرده را پاکسازی میکند، بنابراین میکروگلیا محافظ مغز است.
با این حال، ممکن است وضعیت در اختلالات نورودژنراتیو مانند بیماری آلزایمر متفاوت باشد، شواهدی وجود دارد مبنی بر اینکه میکروگلیا ممکن است بیش از حد فعال شود و التهاب عصبی را تحریک کند که خود منجر به رسوبات پروتئین سمی مشخصه در آلزایمر شود (پلاکهای آمیلوئید و گرههای نوروفیبریلاری). در نهایت، تحقیقات نشان میدهد که میکروگلیا در رشد مغز نقش دارد. به طور معمول، سیناپسها بسیار بیشتر از آنچه لازم است ایجاد میشوند و تنها قویترین و مهمترین آنها باقی میمانند. میکروگلیا با خوردن سیناپسهایی که بهعنوان غیرضروری برچسبگذاری شدهاند، مستقیماً به این فرآیند «هرس» (pruning) سیناپسی کمک میکنند.
ماکروگلیا
این گروه از سلولها از بافت اکتودرمی مشتق شده و شامل موارد زیر هستند:
آستروسیت ها
سلولهای «آستروسیتها» (Astrocytes) سلولهای ستارهای شکل هستند که محیط کار نورون را حفظ میکنند. آنها این کار را با کنترل سطوح انتقال دهندههای عصبی در اطراف سیناپسها، کنترل غلظت یونهای مهم مانند پتاسیم و ارائه پشتیبانی متابولیک انجام میدهند. آستروسیتها به انواع فیبری و پروتوپلاسمی تقسیم میشوند. وظیفه آنها حفظ سد خونی - مغزی و حفظ محیط شیمیایی با بازیافت یونها و انتقالدهندههای عصبی است. از آنجایی که آستروسیتها توانایی درک سطوح انتقالدهندههای عصبی در سیناپسها را دارند و میتوانند با آزاد کردن مولکولهایی که مستقیماً بر فعالیت عصبی تأثیر میگذارند پاسخ دهند، این سلولها به طور فزایندهای برای اصلاح سیناپسها مهم هستند.
الیگودندروسیت ها
«الیگودندروسیتها» (Oligodendrocytes) از آکسونهای نورونها در سیستم عصبی مرکزی پشتیبانی میکنند، به ویژه آنهایی که مسافتهای طولانی را در مغز طی میکنند. آنها مادهای چرب به نام میلین تولید میکنند که به عنوان لایهای از عایق دور آکسونها پیچیده میشود. همانند لایههای عایق اطراف کابلهای برق، غلاف میلین به پیامهای الکتریکی اجازه میدهد تا سریعتر حرکت کنند و به آنها نام به ماده سفید را میدهد، این ماده سفید میلینی دور آکسونها پیچیده شده است. این گروه از سلولها به دو نوع بین فاسیکولار و پیش نورونی تقسیم میشوند و همانطور که بیان شد وظیفه آنها میلینی کردن آکسونها در سیستم عصبی مرکزی و ایجاد یک چارچوب ساختاری کلی است.
سلول های اپاندیمال
«سلولهای اپاندیمال» (Ependymal cells) این سلولها در بطنهای خطی (مغز) و کانال مرکزی (ستون فقرات) حضور داشته و در تولید مایع مغزی - نخاعی نقش دارند.
گلیای شعاعی
«سلولهای گلیال شعاعی» (Radial glia) سلولهای پیشساز هستند که میتوانند نورونها، آستروسیتها و الیگودندروسیتها را تولید کنند.
مواردی که در بالا بررسی شد نوروگلیاهای سیستم عصبی مرکزی بودند اما در سیستم عصبی محیطی نیز ۳ نوع نوروگلیا یا سلول پشتیبان عصبی وجود دارند که شامل موارد زیر هستند:
- «سلولهای شوان» (Schwann cells). این سلولها در میلیندار کردن آکسونها در سیستم عصبی محیطی نقش دارند.
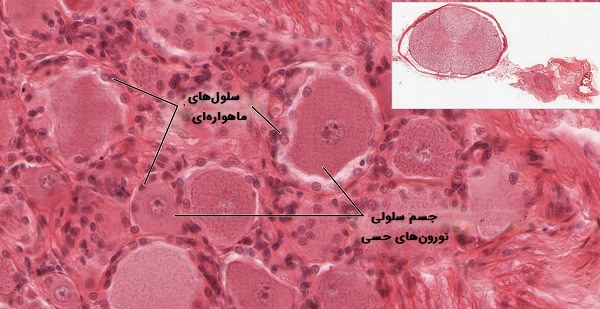
- «سلولهای ماهوارهای» (Satellite cells). سطوح مواد مغذی و انتقال دهندههای عصبی را در اطراف نورونها در گانگلیون تنظیم میکنند.
- «سلولهای گلیال رودهای» (Enteric glial cells). سلولهای گلیال رودهای در اعصاب دستگاه گوارش یافت میشوند.
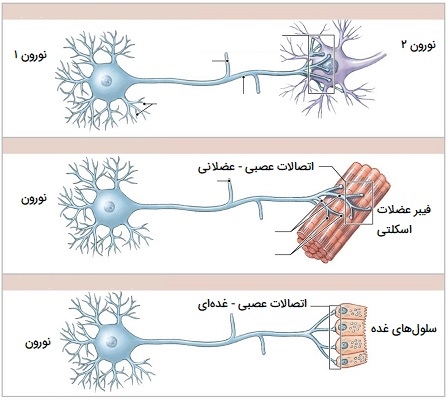
سیناپس چیست؟
بیشتر نورونها سیگنالها را از طریق آکسونهای خود ارسال میکنند، اگرچه برخی از انواع آنها قادر به انتشار سیگنالهایی از دندریتهای خود هستند. در واقع، برخی از انواع نورونها مانند سلولهای آماکرین شبکیه چشم انسان فاقد آکسون هستند و فقط از طریق دندریتهایشان ارتباط برقرار میکنند. سیگنالهای عصبی در امتداد آکسون به شکل امواج الکتروشیمیایی به نام پتانسیل عمل منتشر میشوند که سیگنالهای سلول به سلول را در نقاط تماس به نام سیناپس منتشر میکنند.
انواع سیناپس
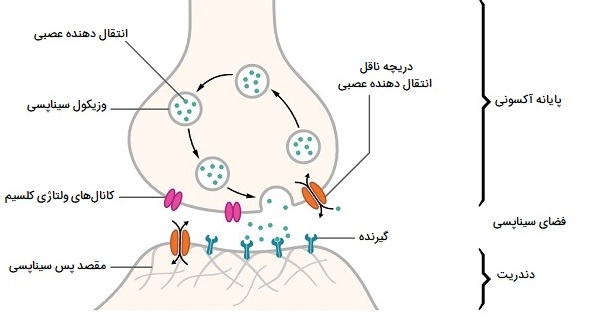
سیناپسها ممکن است الکتریکی یا شیمیایی باشند. سیناپسهای الکتریکی یونها را مستقیماً بین نورونها منتقل میکنند، اما سیناپسهای شیمیایی بسیار رایجتر و از نظر عملکرد بسیار متنوعتر هستند. در یک سیناپس شیمیایی، سلولی که سیگنال میفرستد پیشسیناپسی و سلولی که سیگنال دریافت میکند پسسیناپسی نامیده میشود. هر دو ناحیه تماس پیشسیناپسی و پس سیناپسی پر از ماشین آلات مولکولی هستند که فرآیند سیگنالدهی را انجام میدهند. ناحیه پیشسیناپسی حاوی تعداد زیادی عروق کروی کوچک به نام وزیکولهای سیناپسی است که مملو از مواد شیمیایی انتقالدهنده عصبی هستند.
هنگامی که کلسیم از طریق کانالهای کلسیم دارای ولتاژ وارد ترمینال پیشسیناپسی میشود، آبشاری از مولکولهای تعبیه شده در غشا فعال شده و باعث میشود محتویات برخی از وزیکولها در فضای باریک بین غشای پیشسیناپسی و پسسیناپسی که به آن شکاف سیناپسی میگویند، رها شود. سپس انتقالدهنده عصبی به گیرندههای شیمیایی تعبیه شده در غشای پسسیناپسی متصل میشود و باعث میشود آنها وارد حالت فعال شوند. بسته به نوع گیرنده، تأثیر روی سلول پسسیناپسی ممکن است به روشهای پیچیدهتری تحریککننده، بازدارنده یا تعدیلکننده باشد. به عنوان مثال، آزاد شدن انتقالدهنده عصبی استیلکولین در تماس سیناپسی بین یک نورون حرکتی و یک سلول عضلانی، سلول عضلانی را دپولاریزه (تغییر الکتریکی داخل سلول عضله) کرده و مجموعهای از رویدادها را شروع میکند که منجر به انقباض سلول عضلانی میشود.
کل فرآیند انتقال سیناپسی تنها کسری از میلیثانیه طول میکشد، اگرچه اثرات روی سلول پس سیناپسی ممکن است بسیار طولانیتر باشد (حتی به طور نامحدود، در مواردی که سیگنال سیناپسی منجر به تشکیل یک رد حافظه میشود). به معنای واقعی کلمه حتی در یک گونه واحد صدها نوع مختلف سیناپس وجود دارد. در واقع، بیش از صد ماده شیمیایی ناقل عصبی شناخته شده وجود دارد و بسیاری از آنها انواع مختلفی از گیرندهها را فعال میکنند. بسیاری از سیناپسها از بیش از یک انتقالدهنده عصبی استفاده میکنند، یک ترتیب معمول این است که سیناپس از یک انتقالدهنده عصبی با مولکول کوچک سریع (مانند گلوتامات یا GABA همراه با یک یا چند انتقالدهنده عصبی پپتیدی که نقش تعدیلکننده کندتر را ایفا میکنند) استفاده کند.
کانال های سیناپسی
دانشمندان علوم اعصاب به طور کلی گیرندهها را به دو گروه بزرگ تقسیم میکنند: کانالهای یونی دردار با لیگاند و گیرندههای جفت شده با پروتئین G یا GPCRs که بر سیگنالهای پیامرسان دوم متکی هستند. هنگامی که یک کانال یونی با دروازه لیگاند فعال میشود، کانالی را باز میکند که به انواع خاصی از یونها اجازه میدهد در سراسر غشاء جریان پیدا کنند. بسته به نوع یون، تأثیر روی سلول هدف ممکن است با نزدیک کردن یا دورتر کردن پتانسیل غشایی از آستانه برای تحریک پتانسیل عمل، تحریکی یا بازدارنده باشد. هنگامی که یک GPCR فعال میشود، آبشاری از فعل و انفعالات مولکولی را در داخل سلول هدف آغاز میکند، که در نهایت ممکن است طیف گستردهای از اثرات پیچیده را ایجاد کند، مانند افزایش یا کاهش حساسیت سلول به محرکها، یا حتی تغییر رونویسی ژن.
گیرنده های عصبی
طبق اصل دیل (اصل دیل فقط به نورون پیش سیناپسی اشاره دارد، زیرا پاسخ گیرندههای مختلف پسسیناپسی به یک انتقالدهنده عصبی واحد میتواند در نورونهای مشابه یا مختلف، متفاوت باشد)، که تنها چند استثنا شناخته شده دارد، یک نورون همان انتقالدهندههای عصبی را در تمام سیناپسهای خود آزاد میکند. البته این بدان معنا نیست که یک نورون تأثیر یکسانی را روی همه اهداف خود اعمال میکند، زیرا تأثیر یک سیناپس به انتقالدهنده عصبی بستگی ندارد، بلکه به گیرندههایی که فعال میکند بستگی دارد.
از آنجایی که اهداف مختلف میتوانند (و اغلب انجام میدهند) از انواع مختلفی از گیرندهها استفاده کنند، ممکن است یک نورون اثرات تحریکی بر روی یک مجموعه از سلولهای هدف، اثرات بازدارنده بر روی برخی دیگر و اثرات تعدیلکننده پیچیده روی سایرین داشته باشد. با این وجود، اتفاق میافتد که دو انتقالدهنده عصبی که به طور گسترده مورد استفاده قرار میگیرند، گلوتامات و گاما آمینو بوتیریک اسید (GABA)، هر کدام تا حد زیادی اثرات ثابتی دارند.
گلوتامات انواع مختلفی از گیرندهها را دارد، اما همه آنها تحریککننده یا تعدیلکننده هستند. به طور مشابه، GABA دارای چندین نوع گیرنده گسترده است، اما همه آنها مهاری هستند. چند موقعیت استثنایی وجود دارد که در آنها مشخص شده است که GABA عمدتاً در طول رشد اولیه دارای اثرات تحریکی است. به دلیل این قوام، سلولهای گلوتاماترژیک اغلب به عنوان نورونهای تحریک کننده و سلولهای GABAergic به عنوان نورونهای بازدارنده شناخته میشوند. در واقع این گیرندهها هستند که تحریککننده و بازدارنده هستند، نه نورونها.
انتقال دهنده های عصبی
انتقالدهنده عصبی یک پیامرسان شیمیایی است که از انتهای عصب یک نورون آزاد میشود تا با نورونهای مجاور ارتباط برقرار کند. انتقالدهندههای عصبی توسط نورونهای پیشسیناپسی آزاد میشوند و نورون پسسیناپسی را تحریک یا مهار میکنند. پس از آزاد شدن، انتقالدهندههای عصبی به سرعت در شکاف سیناپسی تجزیه میشوند یا انتقالدهندههای عصبی توسط نورون پیشسیناپسی بازگردانده میشوند (به این حالت بازجذب گفته میشود) تا مدت زمانی که آنها در شکاف سیناپسی هستند محدود شود بنابراین تأثیر انتقالدهنده عصبی محدود میشود. اگر جلو بازجذب یک انتقالدهنده عصبی گرفته شود، تأثیر آن بر نورونهای پس سیناپسی افزایش مییابد. این عملکرد مهارکنندههای بازجذب است، مانند SSRI (مهارکننده انتخابی بازجذب سروتونین) که تأثیر سروتونین بر مغز را افزایش میدهد.
توجه داشته باشید که انتقالدهندههای عصبی به گیرندههای یونوتروپیک متصل میشوند. این گیرندهها کانالهای یونی دردار وابسته به لیگاند (LICs، LGIC)، هستند که معمولاً به عنوان گیرندههای یونوتروپیک نیز شناخته میشوند، گروهی از پروتئینهای کانال یونی گذرنده که به یونهایی مانند +Na+ ،K+ ،Ca2 یا Cl− در پاسخ به اتصال یک پیامرسان شیمیایی (یعنی لیگاند)، مانند یک انتقالدهنده عصبی، اجازه عبور از غشا را میدهند. نمونههایی از انتقالدهندههای عصبی: استیلکولین، دوپامین، گابا، گلایسین، هیستامین، نوراپی نفرین و سروتونین هستند که در ادامه آنها را بیشتر بررسی میکنیم.
- استیلکولین. استیل کولین انتقالدهنده عصبی اصلی سیستم عصبی پاراسمپاتیک، بخشی از سیستم عصبی خودمختار (شاخهای از سیستم عصبی محیطی) است که ماهیچههای صاف را منقبض کرده، رگهای خونی را گشاد میکند، ترشحات بدن را افزایش داده و ضربان قلب را کاهش میدهد.
- دوپامین. دوپامین یک نوع انتقالدهنده عصبی است که بدن آن را میسازد و سیستم عصبی از آن برای ارسال پیام بین سلولهای عصبی استفاده میکند. به همین دلیل است که گاهی اوقات به آن پیامرسان شیمیایی میگویند. دوپامین یک ماده شیمیایی مهم مغز است که بر خلق و خو و احساس پاداش و انگیزه انسان تأثیر میگذارد.
- GABA. گاما آمینوبوتیریک اسید (GABA) یک ماده شیمیایی است که در مغز ساخته شده و در برخی غذاها نیز یافت میشود. در مغز، گابا دارای اثرات ضد تشنج و ضد اضطراب است. علاقه به استفاده از GABA به عنوان یک مکمل غذایی وجود دارد. اما زمانی که گابا از راه خوراکی مصرف شود، ممکن است نتواند از سد خونی مغزی عبور کند. بنابراین مشخص نیست که آیا مکملهای GABA بر مغز تأثیر دارند یا خیر. افراد از GABA برای بسیاری از شرایط مختلف از جمله اختلال استرس و اضطراب، افسردگی و فشار خون بالا استفاده میکنند، اما هیچ مدرک علمی خوبی برای حمایت از استفاده از آن وجود ندارد.
- گلایسین. گلایسین اصلیترین انتقالدهنده عصبی بازدارنده در ساقه مغز و نخاع است که در انواع عملکردهای حرکتی و حسی شرکت میکند. گلایسین چندین عملکرد را به عنوان یک فرستنده در سیستم عصبی مرکزی (CNS) انجام میدهد.
- هیستامین. هیستامین یک انتقالدهنده در سیستم عصبی و یک مولکول سیگنال در روده، پوست و سیستم ایمنی است. هیستامین در نقش خود به عنوان یک انتقالدهنده عصبی، چرخه خواب و بیداری را کنترل میکند. این ماده ممکن است سطح GABA را کاهش دهد و سطح نوراپی نفرین و اپینفرین را افزایش دهد. هیستامین همچنین ممکن است باعث افزایش نفوذپذیری سد خونی - مغزی شود.
- نوراپی نفرین. نوراپی نفرین که نورآدرنالین نیز نامیده میشود، هم هورمونی است که توسط غدد فوق کلیوی تولید شده و نقش پیامرسان شیمیایی را بر عهده دارد و هم در نقش یک انتقالدهنده عصبی، سیگنالها را از انتهای عصبی بدن منتقل میکند. این انتقالدهنده عصبی توسط سیستم عصبی سمپاتیک در انواع تنظیم عصبی استفاده میشود.
- سروتونین. سروتونین یک انتقالدهنده عصبی است که مسئول تنظیم خلق و خو، هضم، خواب و غیره است. سروتونین به عنوان یک انتقالدهنده عصبی به انتقال پیامها از ناحیهای از مغز به ناحیه دیگر کمک میکند.
تعدیل کننده های عصبی
یک «تعدیلکننده عصبی» (Neuromodulator) یک پیامرسان شیمیایی است که از یک نورون در سیستم عصبی مرکزی یا در محیط اطراف آزاد میشود و بر جمعیت متنوعی از نورونها که دارای گیرنده مناسب هستند تأثیر میگذارد. به طور دقیقتر، عملکرد تعدیلکنندههای عصبی برای تغییر قدرت انتقال سیگنال بین نورونها است. تعدیلکنندههای عصبی میتوانند انتقال سیگنال عصبی را با کنترل میزان انتقالدهندههای عصبی سنتز و آزاد شده توسط نورونها تغییر دهند. انتشار تعدیلکنندههای عصبی ممکن است بر روی نورونهای نزدیک محل یا فرایند آزاد شدن تأثیر گذاشته یا ممکن است بر نورونهای بسیار دور از محل انتشار تأثیر بگذارد. این بدان معناست که نورومدولاتورها در مقایسه با انتقالدهندههای عصبی معمولی دارای دامنه عمل بسیار طولانی هستند. به طور کلی میتوان در مورد تعدیلکنندههای عصبی گفت:
- آنها به طور پراکنده از طریق انتقال حجمی آزاد میشوند، یعنی این انتقالدهندههای عصبی به طور کلی در بافت عصبی و نه در یک سیناپس خاص آزاد میشوند، بنابراین بیشتر به عنوان یک سیگنال ارسالی شیمیایی به یک ناحیه (احتمالاً کوچک) مغز عمل میکند تا اینکه نورونهای خاصی را هدف قرار دهد.
- آنها معمولاً از نوع متفاوتی از گیرندههای عصبی استفاده میکنند. انتقالدهندههای عصبی هدفدار و آزادشده از طریق سیناپسی از گیرندههای عصبی یونی سریع الاثر استفاده میکنند که سیگنالهای الکتریکی مثبت (+) و منفی (-) را به نورون هدف منتقل میکنند. با این حال، نورومدولاتورها از گیرندههای عصبی متابوتروپیک یا پروتئین G از سه نوع استفاده میکنند: Gs، Gi، و Gq. اینها گیرندههای کند عمل هستند که عملکرد نورون را در دورههای طولانیتر تنظیم و تعدیل میکنند.
- انتقالدهندههای عصبی خاصی وجود دارند که به عنوان تعدیلکنندههای عصبی طبقهبندی شدهاند، زیرا تقریباً همیشه به روشی که در موارد اول و دوم توضیح داده شده است، کار میکنند. این موارد عبارتند از: دوپامین (D)، سروتونین (5HT)، استیل کولین (M و نیکوتین)، نورآدرنالین (آلفا و بتا)، و هیستامین (H). این انتقالدهندههای عصبی به جای ارسال سیگنالها به نورونهای خاص، عملکرد مدارهای عصبی را در بافت عصبی تنظیم میکنند.
نکته مهم درباره انتقالدهندهها و تعدیلکنندهها این است که نوع گیرندهای که ماده شیمیایی با آن تعامل میکند تعیین میکند که آیا نقش ماده آزادشده انتقالدهنده عصبی است یا تعدیل کننده عصبی. از آنجایی که گیرنده مهم است، همان ماده شیمیایی میتواند هم یک انتقالدهنده عصبی سریع و هم یک تعدیلکننده عصبی کند اثر باشد. در برخی موارد، یک ماده شیمیایی میتواند هر دو اثر را به طور همزمان بر روی یک سلول پس سیناپسی داشته باشد.
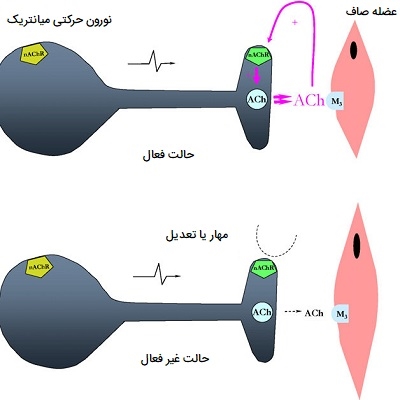
نوروهورمون ها
نوروهورمون یک هورمون عصبی و یک پیام رسان شیمیایی است که توسط سلولهای عصبی غدد ترشح میشود. سلولهای عصبی غدد درونریز سلولهایی هستند که ورودی را از نورونها مانند انتقالدهندههای عصبی دریافت میکنند و در پاسخ، مولکولهای پیامرسان (معروف به هورمونها) را در جریان خون آزاد میکنند. با آزاد کردن هورمونها در گردش خون، هورمونهای عصبی میتوانند اثر خود را روی اهداف محیطی بسیار دور اعمال کنند. نورهورمونها از نظر میزان عملکردشان با تعدیلکنندههای عصبی تفاوت دارند.
نمونهای از سلولهای عصبی غدد درون ریز سلولهای مدولای آدرنال است که درونیترین قسمت غده فوق کلیوی است. مدولای آدرنال در پاسخ به تحریک نورونهای پیشگانگلیونی سمپاتیک، که نورونهای سیستم عصبی خودمختار هستند، آدرنالین را در جریان خون آزاد میکند. همچنین توجه داشته باشید که نوروهورمونها تا 10 برابر بیشتر از انتقالدهندههای عصبی دوام میآورند.
سیستم عصبی چگونه کار می کند؟
سیستم عصبی از تمام سلولهای عصبی بدن شما تشکیل شده است. از طریق سیستم عصبی است که ما با دنیای بیرون ارتباط برقرار میکنیم و در عین حال مکانیسمهای زیادی در داخل بدن ما کنترل میشود. سیستم عصبی از طریق حواس ما اطلاعات را دریافت کرده، آنها را پردازش و واکنشهایی را ایجاد میکند، مانند حرکت دادن ماهیچههای شما یا ایجاد احساس درد. به عنوان مثال، اگر یک بشقاب داغ را لمس کنید، به طور بازتابی دست خود را عقب میکشید و اعصاب شما به طور همزمان سیگنالهای درد را به مغز شما ارسال میکنند. فرآیندهای متابولیک نیز توسط سیستم عصبی کنترل میشود.
میلیاردها سلول عصبی که نورون نیز نامیده میشوند در سیستم عصبی وجود دارد. مغز به تنهایی حدود 100 میلیارد نورون در خود دارد. هر نورون دارای یک جسم سلولی و زائدههای مختلف است. زائدههای کوتاهتر (که دندریت نامیده میشوند) مانند آنتن عمل میکنند: آنها سیگنالهایی را از مثلاً سایر نورونها دریافت میکنند و آنها را به جسم سلولی منتقل میکنند. سپس سیگنالها از طریق یک زائده طولانی (آکسون) که میتواند تا یک متر طول داشته باشد، منتقل میشود.
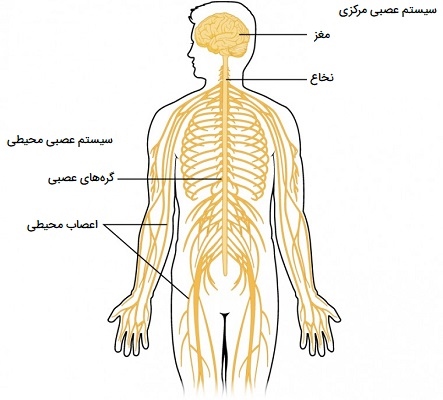
کار سیستم عصبی مرکزی و محیطی
همانطور که پیشتر بیان شد سیستم عصبی دارای دو بخش است که به دلیل قرار گرفتن در بدن، سیستم عصبی مرکزی و سیستم عصبی محیطی نامیده میشود. سیستم عصبی مرکزی (CNS) شامل اعصاب مغز و نخاع است و به طور ایمن در جمجمه و کانال مهرهای ستون فقرات وجود دارند. تمام اعصاب دیگر بدن بخشی از سیستم عصبی محیطی (PNS) هستند. سیستم عصبی مرکزی (CNS) و سیستم عصبی محیطی (PNS) اطلاعات حسی را انتقال داده و پردازش میکنند و عملکردهای بدن را هماهنگ میکنند. مغز و نخاع (CNS) به عنوان مرکز کنترل عمل میکنند. آنها دادهها و بازخورد را از اندامهای حسی و از اعصاب سراسر بدن دریافت میکنند، اطلاعات را پردازش و دستورات را به بیرون ارسال میکنند.
12 جفت اعصاب جمجمهای مغز را به چشمها، گوش انسان و سایر اندامهای حسی و به عضلات سر و گردن متصل میکند. سی و یک جفت اعصاب نخاعی از نخاع به بافتهای قفسه سینه، شکم و اندامها منشعب میشوند. هر عصب مسئول انتقال اطلاعات حسی، ارسال دستورات حرکتی یا هر دو است.
کار نورون ها در اعمال سریع
تمام بافتهای عصبی، از مغز گرفته تا نخاع تا دورترین شاخه عصبی، شامل سلولهایی به نام نورون هستند. نورونها سلولهای باردار هستند: آنها سیگنالهای الکتریکی را برای انتقال اطلاعات از بدن هدایت میکنند. یک نورون معمولی از یک جسم سلولی، دندریتها و یک آکسون با پایانه آکسونی تشکیل شده است. دندریتها سیگنالهایی را از بافتهای بدن یا سایر نورونها دریافت میکنند و آنها را به جسم سلولی منتقل میکنند. اگر سیگنال خروجی تولید شود، آکسون را به سمت پایانه آکسون میپیچد و به نورون یا سلول هدف بعدی میرود. این قابلیت رسانایی اطلاعات را با سرعتی باورنکردنی به بالا و پایین مسیرهای عصبی و از طریق سیستم عصبی مرکزی ارسال میکند. حدود 100 میلیارد نورون به مغز قدرت پردازش فوق العادهای میدهد.
عملکرد انتقال دهنده های عصبی
پیامهای سیستم عصبی به عنوان سیگنالهای الکتریکی از طریق نورونها عبور میکنند. هنگامی که این سیگنالها به انتهای یک نورون میرسند، آزادسازی مواد شیمیایی به نام انتقالدهندههای عصبی را تحریک میکنند. انتقال دهندههای عصبی در سیناپسها، فضاهای بین نورونها با هم یا بین نورونها و سایر بافتها و سلولهای بدن حرکت میکنند. انتقالدهندههای عصبی را میتوان به دو نوع تقسیم کرد: تحریکی یا مهاری. انتقالدهندههای عصبی تحریکی سیگنالهای الکتریکی را در سایر نورونها تحریک کرده و پاسخهای سلولهای بدن را تشویق میکنند. فرستندههای بازدارنده، سیگنالها و پاسخهای سلولی را سست میکنند. از طریق این مواد شیمیایی، سیستم عصبی فعالیت عضلات، غدد و مسیرهای عصبی خود را تنظیم میکند.
نقش طناب نخاعی در انعکاس ها
طناب نخاعی استوانهای کشیده از بدن سلولهای عصبی، دستههای آکسون و سلولهای دیگر است که توسط بافت همبند و استخوان محافظت میشود. در بصلالنخاع به مغز متصل میشود و از ستون مهرهها، تونل توخالی محصور در مهرههای ستون فقرات، میگذرد. نخاع بخشی از سیستم عصبی مرکزی است و به عنوان نوعی بزرگراه عمل میکند. اطلاعات حسی و دستورات حرکتی بالا و پایین رفته و به سمت مغز میروند. این سیگنالها از طریق اعصاب نخاعی به داخل و خارج نخاع شتاب میگیرند. برخی از سیگنالهای دریافتی نیاز به یک پاسخ ساده و فوری دارند. طناب نخاعی میتواند یک فرمان انعکاسی را بدون درگیر کردن مغز انجام دهد.
عملکرد مغز در ادراک و حافظه
سیستم عصبی چیزی بیش از مسیریابی اطلاعات و پردازش دستورات انجام میدهد. چرا بوهای خاص بلافاصله خاطرات خاصی را ایجاد میکنند؟ به نظر میرسد که پاسخ در سیستم لیمبیک نهفته است. سیستم لیمبیک دو حلقه جفتی را در مغز تشکیل میدهد که از هیپوکامپ، آمیگدال، چین سینوسی مغزی کمربندی و شکنج دندانهدار به همراه سایر ساختارها و مجاری تشکیل شده است. همانند سایر بخشهای مغز، سیستم لیمبیک در عملکردها و سطوح مختلف سیستم عصبی نقش دارد. این به پردازش حافظه و بویایی (حس بویایی ما) کمک کرده و طیف وسیعی از احساسات را مدیریت میکند. عطری که از قابلمه روی اجاق بلند میشود ممکن است دست شما را به سمت قاشق دراز کند. همچنین ممکن است یک شام مربوط به زمانهای قبلی شما را بخواند و باعث خوشحالی، نوستالژیک یا یادآوری ناراحتی باشد.
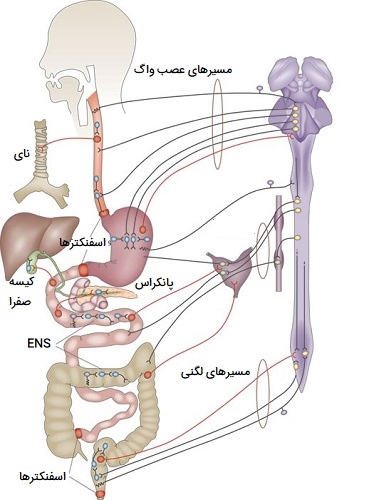
تنظیم عصبی دستگاه گوارش
هضم با مرحله سفالیک (تحریک گیرندهٰهای حسی مانند بویایی و چشایی در سر) شروع میشود که سیستم گوارش را برای دریافت غذا آماده میکند. وقتی مغز ما اطلاعات بصری یا بویایی در مورد غذا را پردازش میکند، تکانههایی را در اعصاب جمجمهای تحریک میکند که غدد بزاقی و معده را برای آماده شدن برای غذا عصبدهی میکند. فاز سفالیک یک پاسخ شرطی یا آموخته شده به غذاهای آشنا است.
اشتها یا میل ما به یک غذای خاص، واکنشهای آمادهسازی مغز را تغییر میدهد. افراد ممکن است در انتظار خوردن پای سیب نسبت به کلم بروکلی، بزاق و صدای معده بیشتری تولید کنند. اشتها و میل محصول هیپوتالاموس و آمیگدال بوده که این نواحی مغز با فرآیندهای احشایی و احساسات مرتبط هستند. پس از مرحله سفالیک، هضم توسط سیستم عصبی روده (ENS) به عنوان یک رفلکس بدون قید و شرط کنترل میشود. افراد مجبور نیستند یاد بگیرند که چگونه غذا را هضم کنند. صرف نظر از اینکه پای سیب باشد یا کلم بروکلی هضم اتفاق میافتد.
سیستم عصبی روده از این نظر منحصر به فرد است که (بیشتر) مستقل از مغز عمل میکند. حدود 90 درصد از ارتباطات پیامهایی هستند که از سیستم عصبی روده به مغز ارسال میشوند تا برعکس. این پیامها به مغز اطلاعاتی در مورد سیری، حالت تهوع یا نفخ میدهد. سیستم عصبی روده، به عنوان بخشی از سیستم عصبی محیطی، همچنین منحصر به فرد است که شامل نورونهای حرکتی و حسی است. به عنوان مثال، سیستم عصبی روده حرکات ماهیچهای صاف را هدایت میکند که غذا را در امتداد دستگاه گوارش، از مری به سمت مقعد هدایت میکند. با این حال، مغز عضلات اسکلتی را هدایت میکند که فرآیندهای آگاهانه مانند بلع و اجابت مزاج را انجام میدهند.
نورونهای حسی سیستم عصبی روده تغییرات معده و روده را تشخیص میدهند. گیرندههای مکانیکی کشش و اتساع پوششهای معده و دوازدهه را زمانی که غذا وارد این حفرهها میشود، تشخیص میدهند. گیرندههای شیمیایی سپس تغییرات در ترکیب شیمیایی کیموس (مایع اسیدی خمیری که از معده به روده میرود) مانند سطح pH و وجود پروتئینها و چربیها را تشخیص میدهند. این اطلاعات برای پیشبرد هر مرحله در هضم و هماهنگی با سیستم غدد درونریز برای ترشح هورمونهای گوارشی استفاده میشود.
اختلالات تنظیم عصبی گوارش
سیستم عصبی روده حاوی 200 تا 600 میلیون نورون است و گاهی اوقات به عنوان مغز کوچک نامیده میشود. از بسیاری از انتقالدهندههای عصبی مشابه سیستم عصبی مرکزی (CNS) و مغز استفاده میکند. به همین دلیل، نورونها در سیستم عصبی روده مستعد اختلالات عصبی مشابه نورونهای مغز هستند و ارتباطی بین اختلالات روده و اختلالات عصبی ایجاد میکنند. به عنوان مثال، در افراد مبتلا به بیماری پارکینسون، نورونها در سیستم عصبی روده و مغز علائم بیماری مشابهی مانند ادخال آلفا سینوکلئین و احتمالاً اجسام لوئی (تودههایی هستند که از تجمع غیرطبیعی پروتئین در داخل سلولهای عصبی تحت تأثیر بیماری پارکینسون ایجاد میشوند) را نشان میدهند. آلفا سینوکلئین (αS) جزء اصلی اجسام لوئی و یک علامت بیماریزای همه سینوکلینوپاتیها از جمله پارکینسون است.
تنظیم عصبی کلیه ها
اعصاب کلیوی حلقه ارتباطی بین سیستم عصبی مرکزی و کلیه هستند. در پاسخ به ورودیهای متعدد محیطی و مرکزی، فعالیت عصب سمپاتیک وابران کلیه تغییر میکند تا اطلاعات را به اجزای اصلی ساختاری و عملکردی کلیه، رگها، گلومرولها و لولهها که هر کدام عصبدهی شدهاند، منتقل کند. در سطح هر یک از این اجزای منفرد انتقال اطلاعات از طریق برهمکنش انتقالدهنده عصبی آزاد شده در محل اتصال پایانه عصب سمپاتیک - عوامل عصبی با گیرندههای پس از اتصال خاص همراه با سیستمهای سیگنالدهی و اثرگذار درونسلولی مشخص انجام میشود.
در پاسخ به محرکهای فیزیولوژیکی طبیعی، تغییرات در فعالیت عصب سمپاتیک وابران کلیه نقش مهمی در تنظیم هومئوستاتیک جریان خون کلیوی، سرعت فیلتراسیون گلومرولی، املاح سلولهای اپیتلیال لولههای کلیوی و انتقال آب و ترشح هورمونی دارد. ورودی آوران از گیرندههای حسی واقع در کلیه در این سیستم کنترل رفلکس از طریق رفلکسهای کلیه شرکت میکند که عملکرد کلی کلیه را قادر میسازد بین دو کلیه خود تنظیم و متعادل شود.
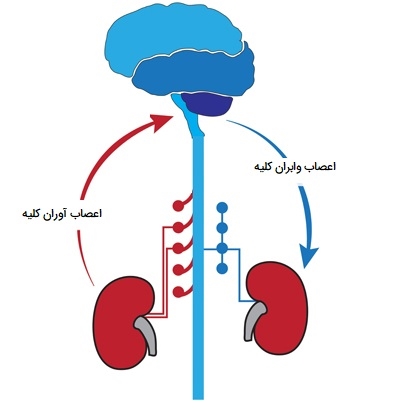
نحوه تنظیم عصبی کلیه و دستگاه ادراری
برخلاف توزیع گسترده رشتههای عصبی سمپاتیک وابران در کلیه، اکثر اعصاب حسی کلیه آوران در ناحیه لگنچه کلیه قرار دارند. اکثر اعصاب کلیه آوران بدون میلین هستند. اعصاب حسی کلیه آوران حاوی ماده P و پپتید مرتبط با ژن کلسیتونین (CGRP) به عنوان انتقالدهندههای عصبی حسی اولیه هستند. وجود اعصاب حسی آوران کلیه در دیواره لگنچه کلیه برای حس کشش دیواره لگنی کلیه ایدهآل است. اجسام سلولی اعصاب کلیوی آوران در گانگلیونهای ریشه پشتی همان طرف (DRG) از T6 تا L4 (شاخههای پشتی اعصاب نخاعی) قرار دارند که در T12 - L3 غالب است.
توزیع اجسام سلولی در بین گونههای مختلف کمی متفاوت است. در طناب نخاعی، اعصاب کلیوی آوران به سمت شاخ پشتی همان طرف در لایههای III ،I تا V جایی که آنها با نورونهای داخلی که به مکانهایی در سیستم عصبی مرکزی مرتبط با تنظیم قلبی عروقی پیش میروند، شامل هسته مجرای منفرد (بخشی از بصلالنخاع و پل مغزی)، بصل النخاع شکمی منقاری (ناحیهای از مغز که مسئول کنترل پایه و رفلکس فعالیت سمپاتیک مرتبط با عملکرد قلبی عروقی است)، اندام زیر فورنیکال (بطن جانبی و سوم مغز را به هم وصل میکند) و هسته پارا بطنی هیپوتالاموس سیناپس میشوند.
همگرایی سیگنالهای آوران از اعصاب سینوس کلیوی و کاروتید بر روی نورونها در چندین ناحیه مغزی که در کنترل قلب و عروق نقش دارند یک مبنای تشریحی برای تعاملات احتمالی بین سیگنالهای آوران ناشی از اندامهای مختلف، از جمله کلیه، فراهم میکند.
تنظیم عصبی پلک زدن
کنترل پلک زدن با ادغام کنترل عصبی خودمختار و به صورت اختیاری هر دو انجام میشود. اعصاب حرکتی در پلکهای فوقانی و تحتانی که توسط اعصاب جمجمه ای VII و III عصبدهی میشوند، باعث انقباض ماهیچههای چشمی و بالابرنده پلک میشوند. بسته شدن پلک به طور خود به خود، با رفلکس یا انقباض ارادی عضلانی رخ میدهد. تصور میشود که کنترل مرکزی پلک زدن خود به خود با فعالیت دمی تنظیم میشود و چندین مطالعه مسیرهای دوپامینرژیک (مسیرهای دوپامینرژیک اتصالات عصبی هستند که در آن دوپامین به مناطقی از مغز و بدن می رود تا اطلاعات مهمی را منتقل کند) را در این کنترل دخیل دانستهاند. فرآیندهای شناختی نیز تأثیر قابل توجهی بر نرخ پلک زدن دارند، به طوری که فعالیتهای فعالتر و ذهنیتری مانند حفظ کردن یا محاسبات ریاضی با افزایش نرخ پلک زدن مرتبط است. نرخ پلک زدن پایین، در مقابل، با دو حالت فیزیولوژیکی مجزا همراه است: بیتوجهی (یا رویاپردازی) و ردیابی محرک، حالتی از دقت بالا که در آن نگاه به یک جسم خاص خیره میشود.
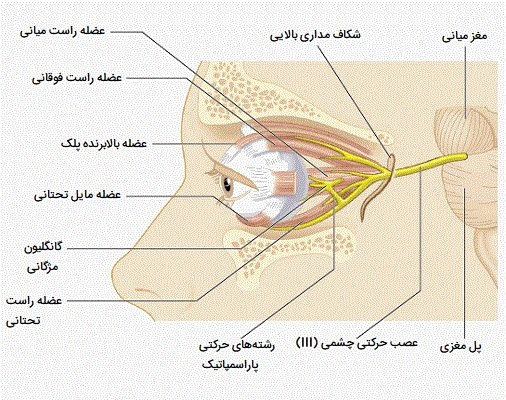
عواملی که پلک زدن را تغییر می دهند
تعدادی از عوامل وجود دارند که میتوانند سرعت پلک زدن را تغییر داده و تأثیر عمیقی بر سطح چشم داشته باشند. نرخ پلک زدن ممکن است بسته به فعالیت در حال تکمیل تغییر کند. نرخ پلک زدن در حالت استراحت بین هشت تا 21 بار در دقیقه تخمین زده میشود، اما اعتقاد بر این است که نرخ پلک زدن در هنگام مکالمه افزایش مییابد و به طور متوسط بین 10/5 تا 32/5 پلک در دقیقه افزایش مییابد. هنگامی که بر روی یک کار بصری خاص تمرکز میکنید، احتمالاً نرخ پلک زدن کاهش مییابد. نشان داده شده است که میزان پلک زدن هنگام مطالعه به طور متوسط به 4/5 پلک در دقیقه کاهش مییابد و افراد در هنگام تغییر خط پلک میزنند.
همچنین نشان داده شده است که فرکانس پلک زدن در هنگام خیره شدن و در حین چشمک زدن تغییر میکند. علاوه بر این، از زمان ظهور رایانههای شخصی در 30 سال پیش، وضعیتی به نام سندرم بینایی رایانه یا CVS ظاهر شده است. یکی از مهمترین مشکلاتی که در CVS نقش دارد، کاهش میزان پلک زدن بیمار است. مطالعات نشان میدهد که افراد هنگام استفاده از رایانه تقریباً 60 درصد کمتر پلک میزنند. این کاهش در سرعت پلک زدن ممکن است نتیجه تمرکز جدی روی کار یا میدان نسبتا محدود حرکت چشم باشد. در نتیجه لایه اشکی بیمار کمتر پر میشود و با سرعت بیشتری تبخیر شده و باعث ناراحتی چشمی او میشود.
میزان پلک زدن نیز به شدت تحت تأثیر عوامل داخلی مانند خستگی، داروهای استرس، احساسات و شرایط سطحی است. در حالی که تأثیر فرآیندهای ذهنی و روند کنترل میزان پلک زدن هنوز تحت بررسی است، به نظر میرسد که نرخ پلک زدن ممکن است توسط عملکرد شناختی و احساسی تغییر کند. به عنوان مثال، افرادی که هیجان عاطفی، اضطراب یا ناامیدی را تجربه میکنند، نرخ پلک زدن افزایش یافته است. همچنین گزارش شده است که احساس گناه بر الگوهای طبیعی پلک زدن تأثیر میگذارد. بیماریهای مرتبط با سطوح غیر طبیعی دوپامین میزان پلک زدن را تغییر میدهند، زیرا اعتقاد بر این است که سطح دوپامین در سیستم عصبی مرکزی با حرکاتی که پلک زدن را کنترل میکنند مرتبط است. بیماران مبتلا به بیماری پارکینسون میزان پلک زدن را کاهش دادهاند، در حالی که بیماران مبتلا به اسکیزوفرنی میزان پلک زدن آنها افزایش یافته است.

داروها همچنین این توانایی را دارند که بر میزان پلک زدن تأثیر بگذارند. زنانی که از قرصهای ضد بارداری استفاده میکنند به طور متوسط 32 درصد بیشتر از کسانی که قرص مصرف نمیکنند پلک میزنند. اگرچه پلک زدن معمولاً افکار آگاهانه زیادی را به همراه نمیآورد، اما بدیهی است که تأثیرات برونزا و درونزا به وضوح نحوه پلک زدن ما را تغییر میدهند.
پلک زدن و تأثیر آن بر بینایی
انجام برخی کارها که معمولاً شامل یک فاصله چشمک زن طولانی مدت میشود میتواند مشکلاتی را برای بیماران مبتلا به اختلال عملکرد سطح چشم ایجاد کند. خشکی و سوزش سطح چشم میتواند توسط این نوع فعالیتها تشدید شود زیرا با نرخ پلک زدن کمتر همراه است. به عنوان مثال، یک کار طولانی مدت بصری مانند کار بر روی رایانه با طولانی شدن فاصله بین پلک زدنها تا 12 ثانیه همراه است. برای بیماران خشکی چشم با زمان شکستن لایه اشک 3 ثانیه، چنین فواصل طولانی احتمالاً منجر به ناراحتی چشمی میشود. با گذشت چند دقیقه، شرایط سطح چشم بدتر میشود و کراتیت (التهاب قرنیه) ممکن است در نتیجه این فواصل متوالی و طولانی بین پلکها افزایش یابد.
تنظیم عصبی تنفس
تهویه حرکت هوا به داخل و خارج از ریه است که تبادل گاز را تسهیل میکند. تنفس از طریق ماهیچههای تنفسی رخ میدهد که بهطور ریتمیک منقبض و شل میشوند تا ریهها را با هوا پر کنند و در هنگام بازدم آنها را تخلیه کنند. در ادامه به کنترل ارادی و غیر ارادی تنفس میپردازیم.
کنترل غیر ارادی تنفس
تنفس غیر ارادی هر نوع کنترل تنفسی است که تحت کنترل مستقیم و آگاهانه نیست. تنفس برای تداوم زندگی لازم است، بنابراین تنفس غیرارادی این امکان را فراهم میکند که در مواقعی مانند هنگام خواب که تنفس ارادی امکانپذیر نباشد فرد اکسیژن لازم را دریافت کند. تنفس غیر ارادی همچنین دارای عملکردهای متابولیکی است که حتی زمانی که فرد هوشیار است کار میکند. تنفس غیر ارادی تحت کنترل ناخودآگاه است. دیافراگم و عضلات بین دندهای، ماهیچههای تنفسی اولیه، توسط گروههایی از نورونهای واقع در پل مغزی و بصلالنخاع تحریک میشوند. این نورونها مرکز کنترل تنفس را تشکیل میدهند. آنها از طریق اعصاب فرنیک و بین دندهای، تکانههایی را به ماهیچههای تنفسی اولیه میفرستند که انقباض آنها را تحریک میکند.
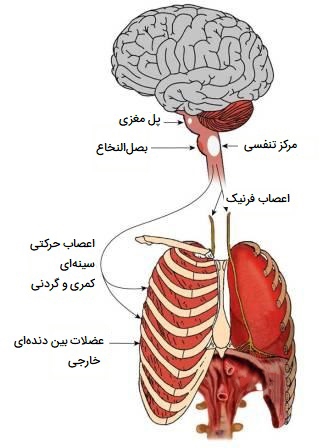
مراکز تنفس غیر ارادی در مغز
تنفس غیر ارادی توسط مراکز تنفسی ساقه فوقانی مغز (گاهی اوقات ساقه تحتانی مغز همراه با مخچه) کنترل میشود. این ناحیه از مغز علاوه بر سیستم تنفسی، بسیاری از عملکردهای غیرارادی و متابولیک، از جمله جنبههای خاصی از عملکرد قلبی عروقی و حرکات غیرارادی عضلات (در مخچه) را کنترل میکند. مراکز تنفسی حاوی گیرندههای شیمیایی هستند که سطوح pH خون را تشخیص میدهند و سیگنالهایی را به مراکز تنفسی مغز میفرستند تا میزان تهویه را برای تغییر اسیدیته با افزایش یا کاهش حذف دیاکسیدکربن (از آنجایی که دی اکسید کربن با سطوح بالاتر یون هیدروژن در خون مرتبط است) تنظیم کنند. همچنین گیرندههای شیمیایی محیطی در سایر رگهای خونی وجود دارد که این عملکرد را نیز انجام میدهند و شامل اجسام آئورت و کاروتید میشود. در ادامه مراکز تنفس غیرارادی در مغز را بیشتر بررسی کردهایم.
بصل النخاع
«بصل النخاع» (The Medulla) مرکز اصلی کنترل تنفسی است. وظیفه اصلی آن ارسال سیگنال به ماهیچههایی است که تنفس را کنترل میکنند تا باعث ایجاد تنفس شود. دو ناحیه در مدولا وجود دارد که تنفس را کنترل میکند:
- گروه تنفسی «شکمی» (ventral): گروه تنفسی شکمی بازدم اجباری ارادی را کنترل کرده و برای افزایش نیروی دم عمل میکند.
- گروه تنفسی «پشتی» (dorsal): گروه تنفسی پشتی (nucleus tractus solitarius) عمدتاً حرکات دمی و زمان آنها را کنترل میکند.
بصل النخاع همچنین رفلکسهای حرکات غیر تنفسی هوا مانند رفلکسهای سرفه و عطسه و همچنین سایر رفلکسها مانند بلع و استفراغ را کنترل میکند.
پل مغزی
«پل مغزی» (The pons) مرکز تنفسی دیگر است و در زیر بصلالنخاع قرار دارد. وظیفه اصلی آن کنترل تعداد یا سرعت تنفس غیرارادی است. دارای دو ناحیه عملکردی اصلی است که این نقش را ایفا میکنند این دو ناحیه شامل موارد زیر هستند:
- مرکز آپنوستیک: این مرکز سیگنالهایی را برای تشویق نفسهای طولانی و عمیق ارسال میکند. شدت تنفس را کنترل کرده و توسط گیرندههای کششی عضلات ریوی در حداکثر عمق دم، یا سیگنالهای مرکز پنوموتاکسیک مهار میشود و همچنین ظرفیت تنفسی را افزایش میدهد.
- مرکز پنوموتاکسیک: این بخش سیگنالهایی را برای مهار دم ارسال میکند که به آن اجازه میدهد تا سرعت تنفس را به خوبی کنترل کند. سیگنالهای آن فعالیت عصب فرنیک را محدود میکند و سیگنالهای مرکز آپنه را مهار میکند و علاوه بر این ظرفیت تنفسی را کاهش میدهد.
سرعت تهویه (حجم دقیقهای) به شدت کنترل میشود و عمدتاً توسط سطح دیاکسیدکربن خون که با سرعت متابولیک تغییر میکند تعیین میشود. گیرندههای شیمیایی میتوانند تغییرات pH خون را که برای تصحیح آن به تغییراتی در تنفس غیرارادی نیاز دارند، تشخیص دهند. بصلالنخاع سیگنالهایی را به ماهیچههایی میفرستد که دم و بازدم را آغاز میکنند و تنظیم عصبی رفلکسهای حرکتی غیر تنفسی هوا مانند سرفه و عطسه را کنترل میکند.
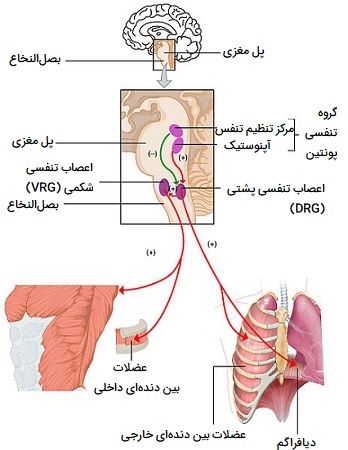
کنترل ارادی تنفس
تنفس ارادی تحت کنترل آگاهانه است. از طریق قشر حرکتی در مغز کنترل میشود که ورودیها را از سیستم لیمبیک و هیپوتالاموس دریافت میکند. مکانیسمهای درگیر در تنظیم عصبی تنفس ارادی به طور کامل شناخته نشدهاند، اما تصور میشود سیگنالهایی از قشر حرکتی به نخاع فرستاده میشوند و سپس به عضلات تنفسی منتقل میشوند.
تنظیم عصبی عطسه
در کلیترین حالت، عطسه با تحریک مخاط بینی در اثر عوامل فیزیکی مانند اجسام خارجی یا مواد شیمیایی مانند مواد تحریککننده شروع میشود. این سیگنال تحریککننده که با آزادسازی هیستامین آغاز شده، توسط انتهای عصب سه قلو جمعآوری میشود و به پیاز نخاعی، مرکز کنترل عطسه که بین مغز و نخاع قرار دارد، منتقل میشود. از پیاز نخاعی دستور عطسه به سیستمهای مختلف صورت، گلو و سینه ارسال میشود. چشمان ما بسته میشود، نفس عمیق میکشیم، گلوت بسته میشود، فشار در ریهها افزایش مییابد و در نهایت گلوت باز میشود و آن خروج تند هوا وجود دارد که عامل ایجادکننده عطسه از طریق بینی و دهان راه خود را به بیرون پیدا میکند.
نتیجه نهایی واکنش عطسه ابری از هزاران قطره کوچک است که دفع آنها فقط 150 میلیثانیه طول میکشد و ضخیمترین قسمت آنها در چند متری میافتد، اما ذرات کوچکتر آنها میتوانند تا هشت متر حرکت کنند و یک اتاق کامل را طی کنند. شدت عطسه به حدی است که مواردی از آسیب به ارگانیسم، تا حد پاره شدن شریان آئورت و حتی بیماری که ایمپلنت دندانی را از بینی خارج کرده است، ثبت شده است. در مورد این واقعیت که وقتی عطسه میکنیم چشمان خود را به طور خودکار میبندیم، باید گفت این به دلیل ارتباط متقابل بین اعصابی که پلک زدن را کنترل میکنند و آنهایی که در عطسه نقش دارند رخ میدهد.
رفلکس عطسه وابسته به نور
به غیر از ذرات تحریک کننده خارجی، آلرژیها یا بیماریهای احتمالی، محرک دیگر قرار گرفتن ناگهانی در معرض نور شدید است، وضعیتی که به عنوان PSR یا رفلکس عطسه نورانی (photic sneeze reflex) شناخته میشود. تمایل به عطسه هنگام قرار گرفتن در معرض نور شدید یک ویژگی اتوزومال غالب است و 18 تا 35 درصد از جمعیت انسان را تحت تأثیر قرار میدهد. بحث های زیادی در مورد علت واقعی و مکانیسم عطسه ناشی از رفلکس عطسه فتاتیک وجود دارد. عطسه در پاسخ به تحریک در حفره بینی رخ میدهد که منجر به انتشار سیگنال فیبر عصبی آوران از طریق شاخههای چشمی و فک بالا عصب سه قلو به هستههای عصب سه قلو در ساقه مغز میشود. این سیگنال در هستههای عصب سه قلو تفسیر میشود و سیگنال فیبر عصبی وابران به قسمتهای مختلف بدن مانند غدد مخاطی و دیافراگم قفسه سینه میرود و در نتیجه عطسه ایجاد میکند.
واضحترین تفاوت بین عطسه طبیعی و عطسه وابسته به نور، محرک آن است که عطسههای طبیعی به دلیل تحریک حفره بینی رخ میدهند، در حالی که عطسه وابسته به نور میتواند ناشی از طیف گستردهای از محرکها باشد. برخی از نظریهها در زیر آمده است. همچنین یک عامل ژنتیکی وجود دارد که احتمال رفلکس عطسه فوتیک را افزایش میدهد. تحریک شاخه چشمی عصب سه قلو ممکن است تحریکپذیری شاخه فک بالا را افزایش داده و در نتیجه احتمال عطسه را افزایش دهد. این شبیه مکانیزمی است که در آن فتوفوبیا با ارسال سیگنالهای مداوم در معرض نور از طریق عصب بینایی و عصب سه قلو ایجاد میشود و باعث افزایش حساسیت در شاخه چشم میشود. اگر این افزایش حساسیت به جای شاخه چشمی در شاخه فک بالا رخ دهد، ممکن است به جای فتوفوبیا عطسه ایجاد شود.

عطسه بعد از غذا خوردن
وضعیتی به نام رینیت چشایی میتواند باعث عطسه برخی افراد بعد از غذا خوردن، به ویژه پس از مصرف غذاهای تند شود. پر شدن معده نمونه دیگری از محرکهایی است که میتواند باعث عطسه غیرقابل کنترل شود. کسانی که این علامت یا اختلال را نشان میدهند که خرناس نامیده میشود، بلافاصله پس از خوردن وعدههای غذایی بزرگ که معده را کاملا پر میکند، بدون توجه به نوع غذای خورده شده، تحت عطسههای غیرقابل کنترل 3 تا 15 عطسه قرار میگیرند. اعتقاد بر این نیست که خرناس یک واکنش آلرژیک از هر نوع باشد.
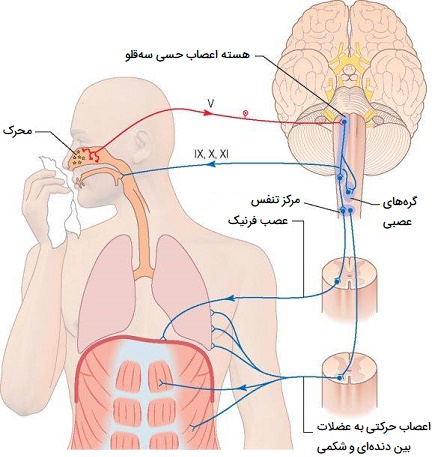
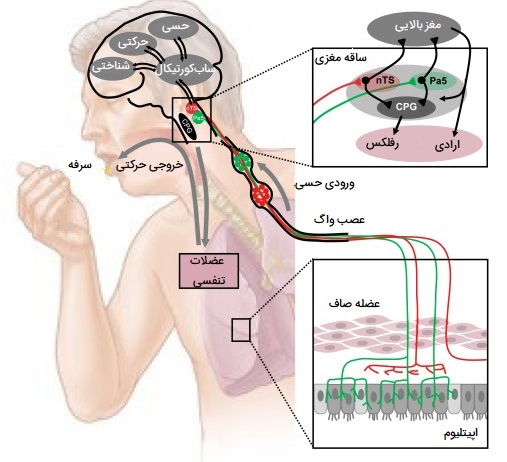
تنظیم عصبی سرفه چگونه است؟
رفلکس سرفه دارای هر دو بخش حسی (آوران) عمدتاً از طریق عصب واگ و اجزای حرکتی (وابران) است. گیرندههای تحریککننده ریوی (گیرندههای سرفه) در اپیتلیوم دستگاه تنفسی به محرکهای مکانیکی و شیمیایی حساس هستند. برونشها و نای به قدری به لمس سبک حساس هستند که مقادیر جزئی ماده خارجی یا سایر علل تحریک رفلکس سرفه را آغاز میکند.
حنجره و کارنیا به ویژه حساس هستند. نایژکهای انتهایی و حتی آلوئولها (کیسههای هوایی) به محرکهای شیمیایی مانند گاز دی اکسید گوگرد یا گاز کلر حساس هستند. هوای با حرکت سریع معمولاً هر ماده خارجی را که در برونش یا نای وجود دارد با خود حمل میکند. تحریک گیرندههای سرفه توسط گرد و غبار یا سایر ذرات خارجی باعث ایجاد سرفه میشود که برای خارج کردن مواد خارجی از مجرای تنفسی قبل از رسیدن به ریهها ضروری است.
فیزیولوژی سرفه
گیرندههای سرفه عمدتاً در دیواره خلفی نای، حلق و در حفره نای، نقطهای که نای به داخل برونش اصلی منشعب میشود، قرار دارند. گیرندهها در مجاری هوایی دیستال کمتر هستند و بالاتر از نایژکهای تنفسی وجود ندارند. هنگامی که تحریک میشوند، تکانهها از طریق عصب حنجره داخلی، شاخهای از عصب حنجره فوقانی که از عصب واگ (CN X) به بصل النخاع مغز منشا میگیرد، حرکت میکند. این مسیر عصبی آوران است. برخلاف سایر نواحی که مسئول اعمال غیرارادی مانند بلع هستند، هیچ ناحیه قطعی که به عنوان مرکز سرفه در مغز شناسایی شده باشد وجود ندارد. سپس مسیر تنظیم عصبی وابران دنبال میشود و سیگنالهای مربوطه از قشر مغز و بصلالنخاع از طریق واگ و اعصاب حنجره فوقانی به گلوت، عضلات بین دندهای خارجی، دیافراگم و سایر عضلات اصلی دمی و بازدمی منتقل میشوند. مکانیسم سرفه به شرح زیر است:
- دیافراگم (عصبرسانی شده توسط عصب فرنیک) و عضلات بین دندهای خارجی (که توسط اعصاب بین دندهای سگمنتال عصبدهی میشوند) منقبض میشوند و فشار منفی در اطراف ریه ایجاد میکنند.
- هوا به سمت ریهها میرود تا فشار را یکسان کند.
- گلوت بسته میشود و تارهای صوتی منقبض میشوند تا حنجره را ببندند.
- عضلات شکم منقبض میشوند تا بر عملکرد دیافراگم آرامشبخش تأکید کنند. به طور همزمان سایر عضلات بازدمی منقبض میشوند. این اعمال باعث افزایش فشار هوا در داخل ریهها میشود.
- تارهای صوتی شل میشوند و گلوت باز میشود و هوا با سرعت بیش از ۱۶۰ کیلومتر بر ساعت خارج میشود.
- نایژهها و بخشهای غیر غضروفی نای فرو میریزند و شکافهایی را ایجاد میکنند که هوا از طریق آن به اجبار وارد میشود و هر گونه محرک چسبیده به پوشش تنفسی را از بین میبرد.
تنظیم عصبی گردش خون چیست؟
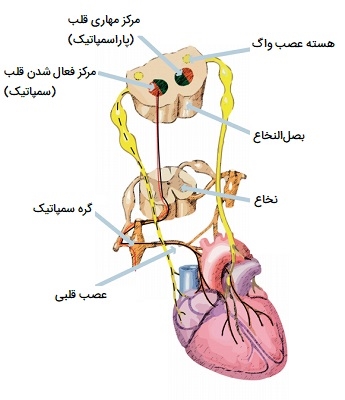
گردش خون محیطی برای توزیع برونده قلبی به اندامها و بافتهای مختلف مطابق با نیازهای متابولیکی یا عملکردی فردی آنها تنظیم میشود و در عین حال فشار خون شریانی را در محدوده نسبتاً باریکی حفظ میکند. مرکز قلب و عروق که در بصل النخاع قرار دارد، شامل سه جزء متمایز است: مرکز شتابدهنده قلب، مرکز بازدارنده قلبی، و مرکز وازوموتور. بدن جریان خون منطقهای را میتواند به طور موثر در سطح محلی توسط توانایی ذاتی عروق برای پاسخ به نیروهای مکانیکی مختلف (مانند کشش دیواره و تنش برشی) و همچنین محرکهای شیمیایی (مانند متابولیتهای بافتی و O2) تنظیم کرد. بر روی این سیستم کنترل محلی، سطح دیگری از تنظیم وجود دارد که توسط تغییرات در فعالیت تنظیم عصبی مرکزی کنترل میشود که عملکرد قلبی عروقی را برای برآوردن نیازهای بدن به عنوان یک کل تنظیم میکند.
مرکز شتابدهنده قلب با تنظیم ضربان قلب و حجم ضربهای از طریق تحریک سمپاتیک عصب شتابدهنده قلب، عملکرد قلب را تحریک میکند. مرکز مهارکنندههای قلبی با کاهش ضربان قلب و حجم ضربهای از طریق تحریک پاراسمپاتیک از عصب واگ، عملکرد قلب را کند میکند. مرکز وازوموتور آهنگ تپشهای عروق یا انقباض عضله صاف در تونیکا را کنترل میکند. تغییر قطر بر مقاومت محیطی، فشار و جریان تأثیر میگذارد که به نوبه خود بر برونده قلبی را تحت تأثیر قرار میدهد. اکثر این نورونها از طریق آزادسازی انتقالدهنده عصبی نوراپینفرین از نورونهای سمپاتیک عمل میکنند. اگرچه هر مرکز به طور مستقل عمل میکند، اما از نظر تشریحی متمایز نیستند.
مرکز قلب و عروق میتواند به محرکهای متعدد پاسخ دهد. هورمونهایی مانند اپینفرین و نوراپینفرین یا تغییرات pH مانند اسیدی شدن به دلیل تجمع دیاکسید کربن در بافت در حین ورزش توسط گیرندههای شیمیایی تشخیص داده میشوند. گیرندههای فشاری که کشش را تشخیص میدهند همچنین میتوانند به مرکز قلبی عروقی سیگنال دهند تا ضربان قلب را تغییر دهد.
تنظیم عصبی فشار خون
تنظیم عصبی فشار خون از طریق نقش مراکز قلبی عروقی و تحریک بارورسپتور به دست میآید. سیستم عصبی اتونومیک (خودمختار) نقش مهمی در تنظیم هومئوستاز عروقی دارد. مکانهای تنظیمی اولیه شامل مراکز قلبی عروقی در مغز است که عملکردهای قلبی و عروقی را کنترل میکنند. تنظیم عصبی فشار خون و جریان آن بستگی به مراکز قلبی عروقی واقع در بصل النخاع دارد. این دسته از نورونها به تغییرات فشار خون و همچنین غلظت اکسیژن، دیاکسیدکربن و سایر عوامل مانند pH در خون پاسخ میدهند.
عملکرد گیرنده های فشاری
بارورسپتورها گیرندههای کششی تخصصی هستند که در نواحی نازک رگهای خونی و حفرههای قلب قرار دارند که به میزان کشش ناشی از حضور خون پاسخ میدهند. آنها برای تنظیم فشار خون، تکانههایی را به مرکز قلب و عروق ارسال میکنند. بارورسپتورهای عروقی عمدتاً در سینوسها (حفرههای کوچک) در داخل آئورت و شریانهای کاروتید یافت میشوند. سینوسهای آئورت در دیوارههای آئورت صعودی درست بالاتر از دریچه آئورت یافت میشوند، در حالی که سینوسهای کاروتید در پایه شریانهای کاروتید داخلی قرار دارند. همچنین گیرندههای فشار کم در دیوارههای ورید اجوف (بزرگ سیاهرگ زبرین) و دهلیز راست قرار دارند.
هنگامی که فشار خون افزایش مییابد، گیرندههای فشاری محکمتر کشیده میشوند و پتانسیل عمل را با سرعت بالاتری آغاز میکنند. در فشار خون پایینتر، میزان کشش کمتر و سرعت شلیک کندتر است. هنگامی که مرکز قلبی - عروقی در بصلالنخاع این ورودی را دریافت میکند، رفلکسی را ایجاد کرده که هومئوستاز را حفظ میکند. هنگامی که فشار خون خیلی بالا میرود، بارورسپتورها با سرعت بیشتری تحریک شده و باعث تحریک پاراسمپاتیک قلب میشوند. در نتیجه برونده قلبی کاهش مییابد. تحریک سمپاتیک شریانهای محیطی نیز کاهش مییابد و منجر به اتساع عروق میشود. در مجموع، این فعالیتها باعث کاهش فشار خون میشوند.

وقتی فشار خون خیلی پایین میآید، سرعت تحریک بارورسپتور کاهش مییابد. این باعث افزایش تحریک سمپاتیک قلب شده و باعث افزایش برونده قلبی میشود. همچنین باعث تحریک سمپاتیک عروق محیطی و در نتیجه انقباض عروق میشود. در مجموع، این فعالیتها باعث افزایش فشار خون میشوند. بارورسپتورهای موجود در ورید اجوف و دهلیز راست فشار خون را کنترل میکنند زیرا خون از گردش خون سیستمیک به قلب باز میگردد. اگر خون با سرعت بیشتری نسبت به خارج شدن از بطن چپ به دهلیز راست بازگردد، گیرندههای دهلیزی مراکز قلبی - عروقی را تحریک میکنند تا تحریک سمپاتیک و برونده قلبی را تا رسیدن به هومئوستاز افزایش دهند. مخالف این حالت هم درست است. این مکانیسم به عنوان رفلکس دهلیزی شناخته میشود.
مکانیسمهای تنظیم عصبی دیگر نیز میتوانند تأثیر قابل توجهی بر عملکرد قلب و عروق داشته باشند. این گونه مکانیسمها شامل سیستم لیمبیک است که پاسخهای فیزیولوژیکی را به محرکهای روانی، رفلکسهای گیرنده شیمیایی، تحریک عمومی سمپاتیک و تحریک پاراسمپاتیک مرتبط میکند.
انواع رفلکس های بدن
در زیستشناسی، یک رفلکس یا عمل بازتابی، یک توالی یا عمل غیرارادی، برنامهریزی نشده و حرکت تقریباً آنی در پاسخ به یک محرک است. یک رفلکس توسط مسیرهای عصبی به نام قوسهای رفلکس امکانپذیر میشود که میتوانند قبل از رسیدن آن ضربه به مغز بر روی یک تکانه عمل کنند. سپس رفلکس یک پاسخ خودکار به محرکی است که فکر آگاهانه دریافت نمیکند یا نیازی به آن ندارد. در ادامه انواع رفلکسهای بدن انسان را توضیح دادهایم.

رفلکس ماهیچه ای
«رفلکسهای کششی میوتاتیک یا عضلانی» (Myotatic reflexes) که گاهی اوقات به عنوان رفلکسهای تاندون عمیق نیز شناخته میشوند اطلاعاتی در مورد یکپارچگی سیستم عصبی مرکزی و سیستم عصبی محیطی ارائه میدهند. به طور کلی، کاهش رفلکسها نشاندهنده یک مشکل محیطی و رفلکسهای پر جنب و جوش یا اغراقآمیز یک مشکل مرکزی است. یک رفلکس کششی انقباض عضله در پاسخ به کشش طولی آن است.
- رفلکس عضله دوسر (C5, C6): پاسخ طبیعی باعث انقباض عضله دو سر بازو میشود و بازو پایینی را به سمت بالا میکشد. راهنمای ارزیابی فیزیکی، این آزمون رفلکس عملکرد قوس بازتابی C5 و قوس بازتابی C6 را بررسی میکند.
- رفلکس براکیورادیالیس (C5, C6, C7): رفلکس براکیورادیالیس با ضربه زدن مستقیم به تاندون براکیورادیالیس با چکش، زمانی که بازوی بیمار در حال استراحت است مشاهده میشود. این نوع رفلکس باید باعث پرونیشن (حرکت به سمت داخل) یا سوپیناسیون (چرخش به خارج) خفیف و خم شدن جزئی آرنج شود.
- رفلکس بازکننده انگشتان (C6, C7): رفلکس بازکننده انگشتان به عنوان بخشی از معاینه عصبی برای ارزیابی مسیرهای حسی و حرکتی در اعصاب نخاعی C6 و C7 آزمایش میشود. همچنین به عنوان رفلکس برونکر - افنبرگ یا BER نیز شناخته میشود.
- رفلکس عضله سه سر (C6, C7, C8): این نوع رفلکس باعث انقباض غیرارادی عضله سهسر بازویی میشود. این رفلکس به عنوان بخشی از معاینه عصبی برای ارزیابی مسیرهای حسی و حرکتی در اعصاب نخاعی C7 و C8 آزمایش میشود.
- رفلکس کشکک یا رفلکس حرکتی زانو (L2, L3, L4): رفلکس کشکک که به آن رفلکس زانو یا حرکت زانو نیز گفته میشود، یک رفلکس کششی است که بخشهای L2، L3 و L4 نخاع را آزمایش میکند. ضربه زدن به تاندون کشکک با یک چکش مخصوص درست در زیر کشکک باعث کشیده شدن دوک عضلانی در عضله چهار سر ران میشود. این ضربه، سیگنالی تولید میکند که به نخاع برمیگردد و (بدون نورونهای داخلی) در سطح L3 یا L4 در نخاع، کاملاً مستقل از مراکز بالاتر سیناپس برقرار میکند. از آنجا، یک نورون حرکتی آلفا یک تکانه وابران را به عضله چهار سر ران هدایت میکند و باعث انقباض میشود. این انقباض، هماهنگ با شل شدن عضله خمکننده همسترینگ متضاد، باعث لگد زدن پا میشود. بین کشش تاندون کشکک و شروع انقباض عضله چهار سر ران، تاخیری در حدود 18 میلیثانیه وجود دارد.
- رفلکس تکان دادن مچ پا (رفلکس آشیل) (S1, S2): رفلکس حرکتی مچ پا که به آن رفلکس آشیل نیز گفته میشود، زمانی رخ میدهد که تاندون آشیل در حالی که پا خمیده پشتی است ضربه میزند. این نوعی رفلکس کششی است که عملکرد عضله گاستروکنمیوس (عضله متصل به تاندون آشیل) و عصب تامینکننده آن را آزمایش میکند. یک نتیجه مثبت تکان دادن پا به سمت سطح کف پا خواهد بود.
رفلکس های مربوط به اعصاب جمجمه ای
انواع رفلکسهای ایجاد شده در بخشهای مختلف جمجمه انسان وجود دارند که در ادامه آنها را بیشتر بررسی میکنیم.
- رفلکس نور مردمک: رفلکس نور مردمک (PLR) یک رفلکس است که قطر مردمک را در پاسخ به شدت (درخشندگی) نوری که بر روی سلولهای گانگلیونی شبکیه در پشت چشم میافتد (در نتیجه به انطباق بینایی با سطوح مختلف روشنایی/تاریکی کمک میکند) را کنترل میکند.
- رفلکس تطابق عدسی چشم: رفلکس تطابق (یا رفلکس تطابق - همگرایی) یک عمل بازتابی چشم است که در پاسخ به تمرکز روی یک جسم نزدیک و سپس نگاه کردن به یک جسم دور (و بالعکس)، رخ میدهد و شامل تغییرات هماهنگ در همگرایی، شکل عدسی و اندازه مردمک است.
- رفلکس تکان دادن فک: رفلکس حرکتی فک یا رفلکس ماستر، یک رفلکس کششی است که برای آزمایش وضعیت عصب سه قلو (عصب جمجمهای V) بیمار و کمک به تشخیص فشردهسازی طناب گردنی فوقانی از ضایعاتی که در بالای فورامن مگنوم هستند استفاده میشود.
- رفلکس قرنیه: همچنین به عنوان رفلکس پلک زدن شناخته میشود. این رفلکس در بالاتر تحت عنوان تنظیم عصبی پلک زدن بررسی شد.
- رفلکس دهلیزی - چشمی: رفلکس دهلیزی چشمی (VOR) یک رفلکس است که برای تثبیت نگاه در حین حرکت سر، با حرکت چشم به دلیل فعال شدن سیستم دهلیزی (بخشی از گوش داخلی درگیر در تعادل) عمل میکند.
- رفلکس گگ: رفلکس حلقی یا رفلکس گگ یک انقباض عضلانی رفلکس پشت گلو است که با لمس سقف دهان، پشت زبان، ناحیه اطراف لوزهها و پشت گلو ایجاد میشود.
رفلکس های نوزادان
نوزادان تازه متولد شده دارای تعدادی رفلکس دیگر هستند که در بزرگسالان دیده نمیشوند که به آنها رفلکسهای اولیه میگویند. این واکنشهای خودکار به محرکها، نوزادان را قادر میسازد تا قبل از هر گونه یادگیری به محیط پاسخ دهند که شامل موارد زیر هستند:
- رفلکس گردن تونیک نامتقارن (ATNR): این رفلکس در انسانهای تازه متولد شده یافت میشود و معمولاً در حدود 6 ماهگی ناپدید میشود. به دلیل موقعیت مشخص بازوها و سر نوزاد که شبیه به قوس شمشیر است، به تیر و کمان یا «بازتاب شمشیربازی» نیز معروف است.
- رفلکس کف دست (PMR): یک رفلکس ابتدایی است که از انقباض عضله چانه با نوازش قسمت خاصی از کف دست ایجاد میشود. در دوران نوزادی وجود دارد و با بالغ شدن مغز در دوران کودکی ناپدید میشود، اما ممکن است به دلیل فرآیندهایی که مسیرهای مهاری طبیعی قشر مغز را مختل میکند، دوباره ظاهر شود.
- رفلکس مورو: رفلکس مورو یک رفلکس نوزادی است که بین هفتههای 28 تا 32 بارداری ایجاد شده و در 3 تا 6 ماهگی ناپدید میشود. این پاسخی است به از دست دادن ناگهانی حمایت و شامل سه جزء مجزا به این ترتیب است: باز کردن بازوها (ربایش)، کشیدن بازوها به داخل (افزایش)، گریه کردن. اهمیت اولیه رفلکس مورو در ارزیابی یکپارچگی سیستم عصبی مرکزی است.
- «رفلکس گرفتن کف دست» (Palmar grasp reflex): رفلکس گرفتن کف دست (یا رفلکس گرفتن) یک رفلکس ابتدایی و غیرارادی است که در نوزادان انسان و اکثر نخستیها یافت میشود. هنگامی که یک جسم، مانند انگشت بزرگسالان، در کف دست نوزاد قرار میگیرد، انگشتان نوزاد به طور انعکاسی آن شی را میگیرند.

- رفلکس مکیدن: رفلکس مکیدن در همه پستانداران مشترک است و در بدو تولد وجود دارد. این امر باعث میشود کودک به طور غریزی هر چیزی را که با سقف دهانش تماس دارد بمکد و نحوه غذا خوردن طبیعی کودک را شبیهسازی کند.
بسیاری از این رفلکسها کاملاً پیچیده هستند و به تعدادی سیناپس در تعدادی از هستههای مختلف در CNS (به عنوان مثال، رفلکس فرار) نیاز دارند. سایر موارد شامل تنها چند سیناپس برای عملکرد هستند. طبق برخی از تعاریف این اصطلاح، فرآیندهایی مانند تنفس، هضم و حفظ ضربان قلب نیز میتوانند به عنوان اعمال رفلکس در نظر گرفته شوند. در پزشکی اغلب از رفلکسها برای ارزیابی سلامت سیستم عصبی استفاده میشود. پزشکان معمولاً فعالیت یک رفلکس را در مقیاسی از 0 تا 4 درجه بندی میکنند.










سلام.
تشکر از مطالب خوب و مفید.
عالی بود این مقاله.
یه سوال داشتم؛
آیا سیستم پاراسمپاتیک و عصب واگ ارتباطی با حملات پنیک اتک دارن؟
با سلام و وقت بخیر،
بله، سیستم عصبی پاراسمپاتیک و عصب واگ در مدیریت حملات پنیک نقش دارند. سیستم پاراسمپاتیک به طور معمول زمانی فعال میشود که بدن در حالت استراحت و آرامش است، بروز حملات پنیک نتیجه فعالیت بیش از اندازه سیستم عصبی سمپاتیک است، بنابراین شاهد بروز علائمی مانند افزایش ضربان قلب، تنفس سریع و احساس وحشت هستیم، در پایان حملات پنیک سیستم عصبی پاراسمپاتیک فعال میشود تا علائم را کاهش دهد و بدن را به حالت عادی بازگرداند.
عصب واگ که از آن نام بردید یکی از عصبهای اصلی سیستم عصبی پاراسمپاتیک است، بنابراین فعالیت آن به بازگرداندن بدن به شرایط طبیعی کمک میکند. در صورتی که تمایل به کسب اطلاعات بیشتر در مورد سیستم عصبی سمپاتیک و پاراسمپاتیک دارید، پیشنهاد میکنیم مطلب «سیستم عصبی سمپاتیک چیست؟ – به زبان ساده» از مجله فرادرس را مطالعه کنید.
از توجه و همراهی شما با مجله فرادرس سپاسگزاریم.
بسیار متشکرم. تخصصی در علوم پزشکی ندارم و تنها از روی علاقه مطالب مرتبط با فیزیولوژی یا آناتومی بدن انسان را دنبال میکنم. با اینحال بقدر ساده و روان و قابل فهم نگارش شده بود که از خواندن آن واقعاً لذت بردم. بازهم تشکر مضاعف.
کامل و جامع و عالی،ممنونم