



بافت ماهیچه ای چیست؟ | ساختار، عملکرد و انواع به زبان ساده
بافت ماهیچه ای یک بافت نرم است که عضلات بدن را تشکیل میدهد و با انقباض خود باعث به حرکت درآمدن استخوان و مفاصل میشود. در این مطلب درباره ساختار، اجزا، عملکرد و انواع بافت ماهیچه ای صحبت کردهایم.
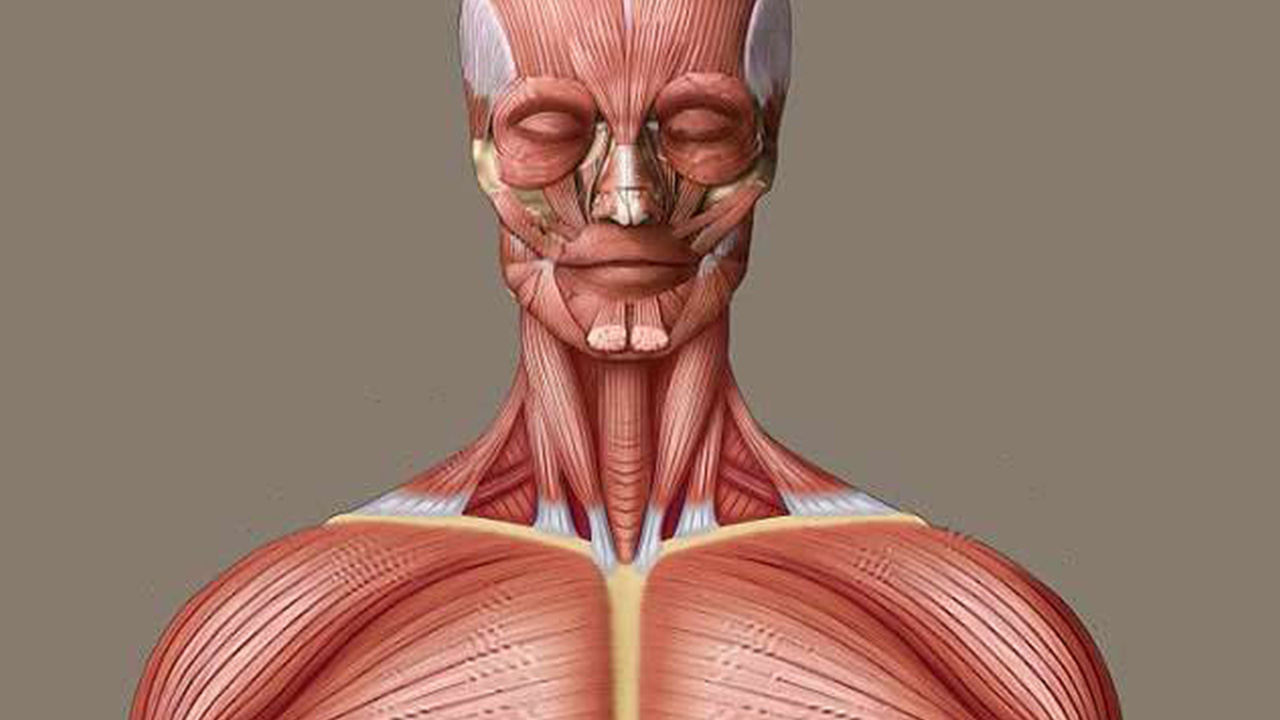


بافت ماهیچه ای چیست؟
بافت ماهیچه ای طی رشد جنین، از طریق فرآیندی شناخته شده به نام میوژنز تشکیل میشود و مسیر تشکیل آن با تاندون و رباطها متفاوت است. بافت ماهیچه ای از سلولهای کشیدهای به نام فیبرهای عضلانی تشکیل میشود. این بافت مسئول حرکات استخوان و مفاصل در بدن و محافظت از ستون مهرهها است. ماهیچهها حاوی پروتئینهای خاصی هستند که با انقباض خود به استخوان و اسکلت نیرو وارد میکنند. انواع بافت ماهیچه ای از نظر عملکرد و موقعیت در بدن متفاوت هستند.
بدن انسان چند عضله دارد؟
بیش از 650 عضله اسکلتی در بدن انسان وجود دارند. سایر بافتهای ماهیچه ای مانند عضلات صاف، به طور معمول در سطح سلولی ظاهر میشوند، به این معنی که میلیاردها سلولِ عضله صاف در بدن وجود دارند که عملکردهای حیاتی مختلفی را انجام میدهند.
بزرگترین ماهیچه های بدن کدام هستند؟
براساس تعاریف مختلف از قدرت قویترین بافتهای ماهیچه ای بدن عبارتند از:
- عضله سرینی (Gluteus Maximus): بزرگترین و قویترین عضله در بدن انسان است چون وظیفه دارد تنه را در حالت ایستاده نگه دارد و عضله اصلی است که به بالا رفتن از پله ها کمک میکند.
- عضلات خارجی چشم (External Muscles of the Eye): عضلات چشم به طور مداوم در حال حرکت هستند تا موقعیت چشم را دوباره تنظیم کنند. هنگامی که سر در حرکت است، عضلات خارجی دائماً موقعیت چشم را تنظیم میکنند تا یک نقطه ثابت شود. با این وجود، عضلات خارجی چشم دچار خستگی میشوند. طی یک ساعت خواندن کتاب، چشمها تقریباً 10 هزار حرکت هماهنگ انجام میدهند.
- عضله قلب: سختترین عضله است. در هر ضربان قلب 71 گرم خون خارج میکند. روزانه قلب حداقل 2500 گالن (9450 لیتر) خون پمپ میکند. قلب توانایی بیش از 3 میلیارد بار پمپاژ در طول زندگی یک فرد را دارد.
- عضله ماضغه یا عضله جونده (Masseter muscle): قویترین عضله بر اساس وزن خود، خصوصا در گیاه خواران است. در حالی که تمام عضلات فک با هم کار میکنند میتواند دندانها را با نیرویی به اندازه 25 کیلوگرم بر روی دندانهای ثناب یا 90/7 کیلوگرم بر روی دندانهای آسیاب فک را ببندد.
- عضلات رحم: رحم در ناحیه تحتانی لگن قرار دارد. این عضلات باید بسیار قدرتمند باشند تا حین فرآیند زایمان، با انقباض آنها نوزاد از کانال زایمان عبور کند. غده هیپوفیز هورمون اکسی توسین ترشح میکند که باعث تحریک انقباضات میشود.
- ماهیچه نعلی یا عضله سولئوس (Soleus): سولئوس در زیر عضله دو قلو قرار دارد و میتواند با بیشترین نیرو به کف پا کشش ایجاد کند. در زیر گاستروکنمیوس (عضله ساق پا) یافت میشود. تکیه برای راه رفتن، دویدن و رقص بسیار مهم است. این عضله در کنار عضلات ساق پا یک عضله بسیار قدرتمند در نظر گرفته میشود زیرا در مقابل نیروی جاذبه بدن را قائم نگه میدارد.
- عضله زبان: زبان از مجموعهای عضلات سخت تشکیل شده است و مانند قلب همیشه کار میکند. این به روند مخلوط کردن مواد غذایی کمک خواهد کرد. خودش را مقید کرده و به شکل حروف در می آورد. زبان حاوی لوزههای زبانی است که میکروبها و پاتوژنها را فیلتر میکند. حتی در هنگام خوابیدن فرد، زبان دائماً بزاق را به سمت گلو هل میدهد.

عضلات مهم بدن کدام هستند؟
به جز مواردی که ذکر شد، برخی دیگر از مهمترین ماهیچههای بدن عبارتند از:
- عضله دلتوئید (Deltoid Muscle): در هریک از شانهها یک ماهیچه دلتوئید قرار دارد. عضلات دلتوئید کمک میکنند تا شانهها به طرفین حرکت کنند.
- عضلات سینه ای (Pectoralis): در هر طرف بالای قفسه سینه دیده میشوند.
- عضلات راست روده شکم (Rectus Abdominus): در زیر سینه، زیر رباط شما یا شکم قرار دارد.
- دو سر بازو (Biceps): اگر بازوهای خود را منقبض کنید این عضلات را حس خواهید کرد.
- عضلات چهار سر ران (Quadriceps): عضلات جلوی ران هستند. دویدن، دوچرخه سواری و ورزشهای پا این عضلات را قدرتمند میکنند.
- ماهیچه جناغی پستانکی (Sternocleidomastoid): عضله اصلی که هنگام چرخش یا کج شدن سر به پهلو و در خم کردن سر به جلو نقش دارد.
قوی ترین ماهیچه بدن کدام است؟
هیچ پاسخی برای این سوال وجود ندارد چون روشهای مختلفی برای اندازهگیری قدرت عضلانی وجود دارند. استحکام مطلق (حداکثر نیرو)، مقاومت دینامیکی (حرکتهای مکرر)، مقاومت الاستیک (سرعت اعمال نیرو) و مقاومت (مقاومت در برابر خستگی) انواع معیارهای تعیین قدرت عضله هستند.
عملکرد بافت ماهیچه ای چیست؟
سلولهای بافت ماهیچه ای تحریکپذیر هستند یعنی به محرک پاسخ میدهند. انقباضی هستند به این معنی که میتوانند با اعمال نیرو کوتاه شوند یا در نبود نیروی فشار، انبساط پیدا کنند. هنگامی که ماهیچه دو جسم متحرک مانند دو استخوان را به هم متصل میکند، انقباض عضلات باعث حرکت استخوانها خواهد شد.
برخی از حرکتهای عضلانی اختیاری هستند به این معنی که تحت کنترل آگاهانه و سیستم اعصاب محیطی بدن قرار دارند. به عنوان مثال، شخصی تصمیم می گیرد کتابی را باز کند و فصلی از آناتومی را بخواند. سایر حرکات مانند انقباض مردمک چشم در برابر نور شدید یا واکنش غیر ارادی نسبت به یک جسم داغ، نا آگاهانه هستند و تحت کنترل اراده قرار ندارند بلکه با سیستم عصبی خودمختار کنترل میشوند.

انواع بافت ماهیچه ای
در پستانداران بافت ماهیچه ای از نظر ساختار و عملکرد به سه دسته تقسیمبندی میشود:
- عضله اسکلتی یا مخطط
- عضله صاف یا نامخطط
- عضله مخطط قلبی یا عضله نیمه مخطط
ماهیچه اسکلتی مخطط
ماهیچه مخطط به استخوان متصل است و انقباض آن باعث ایجاد حرکت، حالت چهره و سایر حرکات ارادی بدن میشود. چهل درصد از توده بدن از عضله اسکلتی تشکیل شده است. عضلات اسکلتی به عنوان محصول جانبی از انقباض خود، گرما تولید میکنند و بنابراین در هومئوستاز حرارتی شرکت دارند. لرزیدن، انقباض غیرارادی عضلات اسکلتی در پاسخ به دمای کمتر از حد طبیعی بدن است. سلول عضلانی یا میوسیت، از میوبلاستهای مشتق شده از مزودرم جنینی ایجاد میشود. میوسیتها و تعداد آنها در طول زندگی به طور نسبی ثابت باقی میمانند.
بافت ماهیچه ای اسکلت به صورت دستههایی احاطه شده توسط بافت همبند چیده شده است. در زیر میکروسکوپ نوری، سلولهای عضلانی به صورت مخروطی دیده میشوند که هستههای فراوان آنها در امتداد غشای سلولی فشرده شدهاند. این رشته به دلیل تناوب منظم پروتئینهای انقباضی اکتین و میوزین همراه با پروتئینهای ساختاری است که پروتئینهای انقباضی را با بافتهای همبند جفت میکند. سلولها در اثر همجوشی میوبلاستهای زیادی که با هم ترکیب شدهاند، فیبرهای عضلانی طویلی را تشکیل میدهند و چند هستهای میشوند.
بافت همبند به عنوان فاشیا (فاسیکل) در تمام عضلات وجود دارد. هر عضله با یک لایه از بافت همبند محاصره شده است که به عنوان اپیمیزیم شناخته میشود. هر فاسيكول را لايهای به نام پريميزيوم و هر فيبر عضله را لایهای از بافت همبند به نام آندوميزيوم احاطه کرده است.

سلول های بافت ماهیچه ای مخطط
ماهیچه مخطط یک الگوی باندی را زیر میکروسکوپ نشان میدهد که ناشی از چیدمان عناصر اسکلت سلولی در سیتوپلاسم فیبرهای عضله است. پروتئینهای سیتوپلاسمی اصلی میوزین و اکتین هستند که در واحدهای تکراری به نام سارکومر منظم شدهاند. اینتراکشن بین میوزین و اکتین مسئول انقباضات ماهیچه ای است. هر اندامک و ماکرومولکول یک فیبر ماهیچه ای طوری قرار گرفته است که امکان برقراری ارتباط و عملکرد بین آنها تضمین شده باشد.
غشای سلولی «سارکولما» و سیتوپلاسم «سارکوپلاسم» گفته میشوند که حاوی میوفیبریلها است. میوفیبریلها پروتئینهای طویلی هستند که شعاع آنها حدود ۱ میکرومتر است و حاوی میوفیلامنت هستند. هسته سلول یا میونوکلئا ساختار مسطحی دارد و از داخل به سارکولما چسبیده است. میتوکندریها بین میوفیبریلها قرار دارند. فیبر عضلانی، شبکه اندوپلاسمی صاف ندارد اما دارای شبکه آندوپلاسمی دانهدار یا خشن است.
اینها از فیبر عضلانی از یک طرف به طرف دیگر عبور میکنند. در بین دو مخزن انتهایی، یک لوله پیچخورده وجود دارد که توبول معکوس (T Tubule) نامیده میشود. این توبولها، مسیرهای بالقوه عمل برای سیگنالسازی شبکه سارکوپلاسمی جهت آزادسازی کلسیم و ایجاد انقباض عضلانی هستند. دو سیسترنای انتهایی (سیسترنا به کیسههای دستگاه گلژی گفته میشود) و یک لوله عرضی همراه با هم، یک سه گانه تشکیل میدهند.
آرایش فیبرهای عضلانی مخطط
معماری عضلانی به آرایش فیبرهای بافت ماهیچه ای اشاره دارد که با محور تولید نیروی عضله مرتبط است. محور در واقع یک خط فرضی کشیده شده از مبدأ ماهیچه به داخل آن است. برای برخی ماهیچههای طولی مانند عضلات دو سرِ بازو، این یک مفهوم ساده محسوب میشود اما برای سایر عضلات مانند عضله راست روده یا عضله دلتوئید، پیچیدگی بیشتری پیدا میکند. در حالی که فیبرهای عضلانی در یک فاسیکول موازی با یکدیگر قرار دارند، خود فاسیکولها میتوانند ارتباط متفاوتی با یکدیگر و تاندونهایشان داشته باشند.
آرایشهای مختلف فیبر، دستههای گستردهای از آرایش عضلات اسکلتی از جمله طولی، پنچهای، تکپایهای، دو پایهای و Multipennate را شامل میشوند. به دلیل وجود این آرایشهای مختلف، کششی که عضله میتواند بین تاندونهای خود ایجاد کند به دلیل چیزی فراتر از اندازه و نوع فیبر آن تفاوت دارد.
آرایش طولی عضله مخطط
فاسیکولهای عضلات طولی، موازی یا دوک مانند، به طور موازی با محور تولید نیرو حرکت میکنند، بنابراین عملکرد آنها روی یک عملکرد کامل شبیه به یک فیبر عضلانی بزرگ و منفرد است. تفاوتهایی وجود دارند که اصطلاحات مختلفی برای آنها استفاده میشوند.
به طور مثال، دوک مانند به آرایش طولی با یک افزایش عرض در قسمت شکمی عضله اشاره دارد مانند عضلات مخطط دو سر بازو، در حالی که آرایش موازی ممکن است یک آرایش طولی روبان مانند مثل عضله راست شکمی باشد. مثالی با رواج کمتر، یک عضله حلقوی مانند ماهیچه حلقوی چشم است که در آن فیبرها طولی مرتب شدهاند اما یک حلقه از مبدأ تا قسمت داخلی تشکیل دادهاند.
آرایش unipennate عضله مخطط
فیبرهای موجود در عضلات Unipennate همه در یک زاویه نسبت به محور تولید نیرو قرار دارند. این زاویه نیروی مؤثر هر فیبر منفرد را کاهش میدهد زیرا در واقع از محور خارج میشود. با این حال به دلیل وجود این زاویه، فیبرهای بیشتری میتوانند در همان حجم بافت ماهیچه ای قرار بگیرند و سطح مقطع فیزیولوژیک (PCSA) را افزایش دهند. این اثر به عنوان بستهبندی فیبر شناخته میشود (از نظر تولید نیرو) و بر افت کارایی، جهت خارج از محور غلبه فراوانی دارد.
این عملکرد با سرعت کوتاه شدن عضلات و در کل روند انجام حرکت انجام میشود. به طور کلی سرعت کوتاه شدن عضلات در مقایسه با سرعت کوتاه شدن فیبر و فاصله کلی کوتاه شدن، کاهش مییابد. تمام این اثرات با زاویه عضلات پری شکل (Pennation) قابل قیاس هستند، زاویههای بزرگتر به دلیل افزایش میزان فیبر و PCSA منجر به نیروی بیشتر میشوند اما با کاهش سرعت بیشتری همراه هستند. «ماهیچه پهن بیرونی» (Vastus Lateralis) نمونهای از معماری Unipennate محسوب میشود.
آرایش Multipennate عضله مخطط
فیبرها در عضلات Multipennate در زوایای مختلفی نسبت به محور تولید نیرو قرار دارند و رایجترین و معمولترین نوع آرایش فیبرهای بافت ماهیچه ای هستند. چندین جهت فیبر شامل دو پایه، همگرا و چند منظوره در این گروه قرار میگیرند. در حالی که تعیین PCSA در این ساختارهای عضلانی دشوارتر میشود، اما همان مبادلات ذکر شده در بالا اعمال میشود. آرایشهای Bipennate اساساً فیبرهایی «V» شکل هستند (مانند ماهیچه راست رانی) که روی هم قرار گرفتهاند.
آرایش Pennation عضله مخطط
آرایشهای همگرای مثلثی یا پری شکل هستند، ریشههای گسترده و الحاقات باریکتری دارند. انواع گسترده زوایای عضلات پری شکل در این آرایش میتوانند چندین عملکرد داشته باشند. به عنوان مثال، عضله ذوزنقهای، یک عضله همگرا نمونه اولیه است که میتواند به اتساع یا جمع شدن شانه کمک کند.
آرایش Multipennate عضله مخطط
آرایشهای Multipennate محدود به یک ترتیب خاص در فیبرها نیستند بلکه به طور خاص به آنچه در اصل ترکیبی از آرایشهای Bipennate یا Unipennate با آرایشهای همگرا است. نمونهای از این معماری میتواند عضله دلتایی باشد.

انواع ماهیچه اسکلتی بر اساس عملکرد
بسیاری از انواع بافت ماهیچه ای بر اساس عملکرد خود نامگذاری میشوند که عبارتند از:
- «خم کننده» (Flexor): باعث کاهش زاویه قدامی در مفصل میشود.
- «اکستنسور» (Extensor): زاویه قدامی را در مفصل افزایش می دهد.
- «دور کننده» (Abductor): استخوان را از خط میانی دور میکند.
- «نزدیک کننده» (Adductor): استخوان را به خط میانی نزدیک میکند.
- «بالابر» (Levator): ساختار را به سمت بالا میبرد.
- «منقبض شونده یا کاهنده» (Depressor): ساختار متصل به خود را به سمت پایین میبرد.
- «سوپیناتور» (Supinator): کف دست را به سمت بالا میچرخاند.
- «درونگَردان» (Pronator): در بخش ساعد قرار دارد و کف دست را به سمت پایین حرکت میدهد.
- «اسفنگتر» (Sphincter): اندازه دهانه را کاهش میدهد.
- «تنسور» (Tensor): بخشی از بدن را منقبض میکند.
- «چرخاننده» (Rotator): استخوان را به دور محور خود میچرخاند.
دستگاه انقباضی عضله اسکلتی
ارتباط فضایی بین رشتههایی که میوفیبریلها را در رشتههای عضلانی اسکلتی تشکیل میدهند بسیار منظم است. این سازمان منظم از عضلات عضلانی باعث ایجاد تقارن عرضی میشود که عضلات اسکلتی و قلبی را مشخص میکند. مجموعههای استریا منفرد مربوط به کوچکترین واحد انقباضی عضله اسکلتی، سارکومر است.

ردیفهای سارکومر، میوفیبریل را تشکیل میدهند که در طول فیبر عضلانی اسکلتی گسترش مییابد. بسته به توزیع و اتصال میو رشتهها، تعدادی از باندها و خطوط را میتوان در سارکومرها تشخیص داد:
- باند I: (رشتههای اکتین، باند A) رشتههای میوزین که ممکن است با رشتههای اکتین همپوشانی داشته باشند.
- باند H: تنها منطقه رشتههای میوزین (بدون همپوشانی با رشتههای اکتین) در باند A
- Z-line: منطقه کاربرد رشتههای اکتین متعلق به دو سارکومر همسایه (با واسطه پروتئینی به نام آلفا اکتینین)
- خط M: باند اتصالات بین رشتههای میوزین (با واسطه پروتئینها، به عنوان مثال میومسین، پروتئین M) هستند.
طول متوسط هر سارکومر حدود 2/5 میکرومتر است (1/5 میکرومتر منقبض شده، 3 میکرومتر کشیده شده). پروتئین تیتین از خط Z به خط M گسترش مییابد و به رشته Z و رشتههای میوزین متصل میشود. تیتین دارای یک قسمت الاستیک است که بین خط Z و مرز بین باند I و A قرار دارد. تیتین به حفظ تراز رشتههای دستگاه انقباضی و مقاومت در برابر کشش منفعل فیبرهای عضلانی کمک میکند. سایر پروتئینهای اسکلتی سلولی خطوط Z میوفیبریلهای همسایه را به هم متصل میکنند. به دلیل این اتصال، باندهای A و I میوفیبریلهای همسایه در فیبر عضلانی کنار هم قرار میگیرند. این پروتئینهای اسکلتی همچنین خطوط Z میوفیبریل محیطی را به سارکولما متصل میکنند.

انواع بافت ماهیچه ای اسکلتی
نوع فیبر با الگوی تحریک فیبر و به نوبه خود با توجه به نوع نورونی که به عضله عصبرسانی میکند، تعیین میشود. اگر الگوی تحریک به صورت آزمایشی تغییر کند، نوع فیبر نیز متناسب با آن تغییر خواهد کرد و این از اهمیت بالینی و پاتولوژیک برخوردار است. فیبرهای عصبی توانایی تشکیل شاخههای جدید به عنوان مثال جوانه زدن و عصبکشی مجدد فیبرهای عضلانی را دارند که ممکن است در اثر ضایعه حاد عصب یا اختلال تخریب عصبی، عصب دهی خود را از دست داده باشند. اگر نوع تحریک حاصل از فیبر عصبی جوانه زده با نوع عضله مطابقت نداشته باشد، نوع فیبر عضلانی تغییر خواهد کرد. روند بازسازی و تنظیم نوع ممکن است منجر به گروه بندی نوع فیبر در عضله شود، یعنی مناطق وسیعی از عضله توسط فیبرهای عضلانی مشابه هم، میشوند.
انواع فیبرهای عضله اسکلتی
انواع مختلفی از فیبرها در بافت ماهیچه ای از نوع اسکلتی وجود دارند که عبارتند از:
- نوع I
- نوع IC
- نوع IIC
- نوع IIAC
- نوع IIA
- نوع IIX
این فیبرها فیبرها بر اساس سرعت منقبض شدن سریع یا آهسته تعریف و در سه نوع اصلی فیبر قرمز، فیبر سفید و فیبر مرکب طبقهبندی میشوند.
فیبر قرمز عضله اسکلتی
ماهیچههای قرمز یا فیبر نوع I، حاوی سلولهای قرمز عضلانی هستند. فیبرهای عضلانی قرمز به نسبت نازک و حاوی مقادیر زیادی میوگلوبین و میتوکندری هستند. فیبرهای قرمز حاوی ایزوفرم میوزین با فعالیت آنزیم ATPase کم است، یعنی سرعتی که میوزین از ATP استفاده میکند بنابراین انقباض کند است. عضلات قرمز در مواقعی که تولید مداوم نیرو ضرورت دارد به عنوان مثال در کنترل وضعیت بدن، استفاده میشود.
فیبر سفید عضله اسکلتی
سلولهای عضلانی سفید یا فیبر نوع II، که به طور عمده در ماهیچههای سفید یافت میشوند، ضخیمتر هستند و میوگلوبین کمتری دارند. فعالیت ATPase ایزوفرم میوزین در فیبر سفید زیاد و انقباض سریع است.
- فیبر نوع IIA (قرمز): حاوی میتوکندریهای زیادی هستند و هم برای فعالیت مداوم و هم برای انقباضات شدید و کوتاه مدت در دسترس هستند. فیبر اکسیداتیو با انقباض سریع (FO)، واسطهای بین فیبر اکسیداتیو کند و فیبر گلیکولیتیک سریع است. این نوع را فیبر را گاهی اوقات فیبر میانی مینامند زیرا دارای خصوصیاتی بین فیبرهای سریع و فیبر کند هستند.
- فیبر نوع IIB/IIX (سفید): حاوی میتوکندریهای کمی هستند. در صورت شتاب سریع و انقباض، به صورت کوتاه مدت فعال میشوند. فیبر نوع IIB/IIX برای تولید ATP مورد نیاز خود و انقباض، به گلیکولیز بیهوازی متکی هستند. گلیکولیتیک سریع انقباض سریع (FG) - در درجه اول از گلیکولیز بیهوازی استفاده میکند. فیبرها بر اساس سرعت منقبض شدن سریع یا آهسته تعریف میشوند. از الیاف برای تولید سریع انقباض اجباری و انجام حرکات سریع و قدرتمند استفاده میشود. با این حال، این فیبرهای عضلانی به سرعت خسته میشوند و در کوتاه مدت فعال هستند. عملکرد این دسته از بافت ماهیچه ای تا حد زیادی به ژنتیک بستگی دارد.

دوک های عضلانی
دوکهای عضلانی تخصص حسی بافت عضلانی هستند. تعدادی از فیبرهای عضلانی کوچک درون فوزلی (فیبر کیسه هستهای و فیبر زنجیره هستهای) توسط یک کپسول بافت همبند احاطه شدهاند. «فیبرهای داخل دوکی» (Intrafusal Muscle Fibers) توسط رشتههای عصبی حرکتی وابران عصبی میشوند. رشته های عصبی حسی آوران فیبرهای عضلانی داخل رحمی را احاطه میکنند. اگر عضله کشیده شود، رشتههای عضلانی در اسپیندل عضله کشیده میشوند، اعصاب حسی تحریک خواهند شد و تغییر در انقباض عضله درک میشود.
انواع مختلف فیبرهای داخل مغزی و انتهای عصب درک موقعیت، سرعت و تسریع انقباض عضله را امکان پذیر میکند. انقباض رشتههای داخل مغزی پس از تحریک توسط رشتههای عصبی وابران، ممکن است تغییرات تحمیل شده بر روی دوک عضله توسط عضله اطراف را خنثی یا بیشتر کند. فیبرهای داخل رحمی و اعصاب وابران از این طریق میتوانند حساسیت را برای عصب حسی که در دوک عضله ختم میشود، تنظیم کنند.
عضله مخطط قلبی
عضله قلبی یا کاردیومیوسیت، دیوارههای انقباضی قلب را تشکیل میدهد. سلولهای عضله قلب معروف به کاردیومیوسیتها، زیر میکروسکوپ به صورت مخطط دیده میشوند. برخلاف بافت ماهیچه ای اسکلتی، کاردیومیوسیتها سلولهای منفرد با یک هسته مستقر در مرکز هستند. یک ویژگی اصلی در کاردیومیوسیتها این است که آنها بر اساس ریتم ذاتی خود بدون تحریک خارجی منقبض میشوند.
کاردیومیوسیتها با اتصالات سلولی ویژهای به نام دیسکهای مقطعی به یکدیگر متصل میشوند. دیسکهای مقطعی دارای اتصالات لنگری و اتصالات شکافدار هستند. سلولهای متصل، فیبرهای عضلانی قلبی منشعب و طولانی را تشکیل میدهند که به عنوان یک هم سنس عمل میکنند و به سلولها اجازه میدهند اقدامات خود را همزمان کنند. عضله قلب، خون را از طریق بدن پمپاژ میکند و تحت کنترل غیر ارادی قرار دارد.

بافت ماهیچه ای قلب زیر میکروسکوپ
با مشاهده این بافت زیر میکروسکوپ، میتوان عضله قلب را به دیواره یک خانه تشبیه کرد. بیشتر دیواره توسط آجرها گرفته میشود که در عضله قلب سلولهای بافت ماهیچه ای قلب یا سلولهای قلبی هستند. ملاتی که آجرها را احاطه کرده است به عنوان ماتریس خارج سلولی شناخته میشود که توسط سلولهای پشتیبانی کننده معروف به فیبروبلاست تولید میشوند.
به همان روشی که دیوارهای خانه شامل سیمهای برق و لولهکشی هستند، عضله قلب نیز دارای سلولهای ویژهای برای انتقال سریع سیگنالهای الکتریکی (سیستم هدایت قلبی) و عروق خونی برای رساندن مواد مغذی به سلولهای عضلانی و دفع مواد زائد است (عروق کرونر، رگها و شبکه مویرگی).
سلول های بافت عضله قلبی
سلولهای عضلانی قلب یا کاردیومیوسیتها سلولهای انقباضی هستند که به قلب اجازه پمپاژ میدهند. هریک از کاردیومیوسیتها باید با هماهنگی سلولهای همسایه خود منقبض شود که به عنوان یک سینسیسیوم عملکردی شناخته میشود و برای پمپاژ مؤثر خون از قلب عمل میکند. اگر این هماهنگی از بین برود (با وجود انقباض سلولهای جداگانه) ممکن است قلب اصلاً پمپاژ نکند، مثل اتفاقی که هنگام ریتمهای غیر طبیعی قلب (آریتمی) در مشکلاتی مانند فیبریلاسیون بطنی رخ میدهد.
سلولهای عضلانی قلب زیر میکروسکوپ تقریباً مستطیل شکل هستند و اندازه آنها 100 تا ۱۵۰ میکرومتر در 30 الی ۴۰ میکرومتر است. این سلولها در انتهای خود توسط دیسکهای مقطعی به هم متصل میشوند و فیبرهای طولانی ایجاد میکنند. هر سلول این بافت ماهیچه ای حاوی میوفیبریلها، فیبرهای پروتئینی ویژهای است که از کنار هم میلغزند. این سلولها به صورت سارکومر، واحد اصلی انقباضی سلولهای بافت ماهیچه ای سازمان یافتهاند.
سازماندهی منظم میوفیبریلها به صورت سارکومر به سلولهای بافت ماهیچه ای قلب زیر میکروسکوپ، شبیه به عضله اسکلتی، حالت راه راه یا مخطط دارد. این رشتهها توسط باندهای سبکتر I ایجاد شدهاند عمدتا از پروتئینی به نام اکتین تشکیل میشوند و باندهای A تیرهتر هستند و عمدتا از میوزین تشکیل شدهاند.
کاردیومیوسیتها حاوی توبولهای T، کیسههای غشایی هستند که از سطح به داخل یاخته منتهی میشوند و به بهبود کارایی انقباض کمک میکنند. اکثر این سلولها فقط یک هسته دارند (اگرچه ممکن است تا چهار هسته هم داشته باشند)، برخلاف سلولهای عضلانی اسکلتی که به طور معمول حاوی هستههای زیادی هستند. سلولهای عضلانی قلب حاوی میتوکندریهای فراوانی هستند که انرژی مورد نیاز سلول را به شکل آدنوزین تری فسفات (ATP) تأمین میکنند و باعث میشوند آنها در برابر خستگی بسیار مقاوم باشند.
توبول های T
توبولهای T لولههای میکروسکوپی هستند که از سطح سلول به عمق سلول منتهی میشوند و با غشا سلولی در یک راستا قرار دارند بوده و از همان دو لایه فسفولیپیدی تشکیل شدهاند و در سطح سلول به مایع خارج سلول راه دارند. توبولهای T در عضله قلب بزرگتر و گستردهتر از عضلات اسکلتی هستند اما تعداد آنها کمتر است. در مرکز سلول به یکدیگر متصل میشوند و به عنوان یک شبکه محوری عرضی به داخل سلول میروند.
در داخل سلول نزدیک به ذخیره داخلی کلسیم سلول، شبکه سارکوپلاسمی قرار دارند. در اینجا، یک لوله واحد با بخشی از شبکه سارکوپلاسمی جفت میشود که در آن، ترکیب دیال نام دارد. عملکرد توبولهای T شامل انتقال سریع تکانههای الکتریکی یا عنوان پتانسیلهای عمل، از سطح سلول به هسته و کمک به تنظیم غلظت کلسیم در سلول است که طی فرآیند اتصال تحریک - انقباض شناخته انجام میشود.

دیسک های مقطعی
سنسیسیوم قلبی شبکهای از کاردیومیوسیتها است که توسط دیسکهای مقاطعی به هم متصل شدهاند و انتقال سریع تکانههای الکتریکی را از طریق شبکه امکانپذیر میکنند و باعث میشوند که سنسیسیوم بتواند در یک انقباض هماهنگ میوکارد عمل کند. یک سنسیسیوم دهلیزی و یک سنسیسیوم بطنی وجود دارد که توسط فیبرهای اتصال قلب متصل میشوند. مقاومت الکتریکی از طریق دیسکهای مقطعی بسیار کم است، بنابراین اجازه میدهد انتشار یون آزاد باشد.
سهولت حرکت یون در امتداد محور فیبرهای عضلانی قلب، به حدی است که پتانسیل عمل قادر به حرکت از یک سلول عضلانی قلب به سلول دیگر است و تنها با مقاومت جزئی مواجه هستند. هر سنسیسیوم از قانون همه یا هیچ پیروی میکند.
عضله صاف
انقباض بافت عضله صاف مسئول حرکات غیر ارادی اندامهای داخلی است. بافت ماهیچه ای عضلات صاف جزئی از سیستم انقباضی دستگاه گوارش، ادراری، تولید مثل و همچنین مجاری تنفسی و عروق خونی هستند. هر سلول عضله صاف، به صورت دوکی و با یک هسته واحد و بدون هیچگونه خطوطی قابل مشاهده است. عضله صاف به دو دسته تقسیم میشود:
- عضله صاف تک واحدی: درون این سلولها کل باند یا صفحه عضلانی به عنوان یک سینسیتیوم منقبض میشود.
- عضله صاف چند واحدی: هر واحد به صورت جداگانه منقبض میشود.

ساختار مولکولی عضله صاف
بخش قابل توجهی از حجم سیتوپلاسم سلولهای عضلات صاف توسط مولکولهای میوزین و اکتین پر شده است که قابلیت انقباض دارند و از طریق یک زنجیره از ساختارهای کششی، کل بافت عضله صاف با آنها منقبض میشود.
میوزین
میوزین در درجه اول عضلات صاف کلاس II است. میوزین II شامل دو زنجیره سنگین (MHC) است که دامنههای سر و دم را تشکیل میدهند. هر یک از این زنجیرههای سنگین، حاوی سر N ترمینال هستند، در حالی که دم در انتهای C، یک مورفولوژی سیم پیچدار به خود میگیرد و دو زنجیره سنگین را به هم میچسباند (تصور کنید دو مار به دور یکدیگر پیچیده شدهاند). بنابراین، میوزین II دارای دو سر است.
در عضلات صاف، یک ژن منفرد (MYH11) زنجیرههای سنگین میوزین II را کد میکند اما انواع مختلف اتصال این ژن وجود دارد که منجر به تولید چهار ایزوفرم میشود. همچنین، عضله صاف ممکن است حاوی MHC باشد که در انقباض بافت ماهیچه ای نقش ندارد و میتواند از ژنهای متعددی ساخته شود. میوزین کلاس دو، ۴ زنجیره سبک هم دارد (MLC) که دارای ۲ سر هستند و وزن آنها ۲۰ (MLC20) و ۱۷ (MLC17) کیلودالتون است.
- MLC20: به عنوان زنجیره سبک تنظیم کننده هم شناخته میشود و به طور فعال در انقباض عضله شرکت میکند. MLC20 دو ایزوفرم در عضلات صاف دارد که توسط ژنهای مختلفی کدگذاری شدهاند اما فقط یک ایزوفرم در انقباض شرکت میکند.
- MLC17: به نام زنجیره سبک ضروری شناخته میشود. عملکرد دقیق آن مشخص نیست اما اعتقاد بر این است که همراه با MLC20، به ثبات ساختاری سر میوزین کمک میکند. دو نوع MLC17 یعنی MLC17a و MLC17b، بر اثر اتصال متناوب در ژن MLC17 وجود دارند.
ترکیبات مختلف زنجیرههای سنگین و سبک میوزین امکان ایجاد صدها نوع مختلف ساختار میوزین را فراهم میکنند اما بعید به نظر میرسد که بیش از چند ترکیب در واقع در یک بستر عضلانی صاف خاص استفاده شده باشند. در رحم، تغییر در بیان میوزین برای تغییر در جهت انقباضات رحمی طی چرخه قاعدگی دیده میشود.

اکتین
رشتههای نازکی که بخشی از دستگاه انقباضی هستند و عمدتا از آلفا اکتین و گاما اکتین تشکیل شدهاند. آلفا اکتین ایزوفرم غالب در عضله صاف است. همچنین مقدار زیادی از اکتین (عمدتا بتا اکتین) وجود دارند که در انقباض شرکت نمیکنند اما در حضور یک محرک انقباضی زیر غشای پلاسما پلیمریزه میشوند و ممکن است به کشش مکانیکی کمک کنند. آلفا اکتین به عنوان ایزوفرمهای ژنتیکی متمایزی مانند آلفا اکتین عضله صاف، عضله قلبی و ایزوفرمهای خاص عضله اسکلتی بیان میشود.
سایر پروتئین های دستگاه انقباضی
عضله صاف حاوی پروتئین تروپونین نیست. در عوض کالمودولین (که نقش تنظیم کننده دارد)، کالدسمون و کالپونین، پروتئینهای مهمی هستند که در بافت ماهیچه ای صاف بیان میشوند که ممکن است هر سه در مهار فعالیت ATPase کمپلکس میوزین نقش داشته باشند که در غیر این صورت انرژی لازم را برای سوخت انقباض عضله فراهم میکند.
- «تروپومیوزین» (Tropomyosin): در عضله صاف وجود دارد، هفت مونومر اکتین را در بر میگیرد و در انتهای طول رشتههای نازک قرار دارد. در عضلات مخطط، تروپومیوزین تا زمانی که کلسیم وجود داشته باشد، مانع از فعل و انفعالات اکتین - میوزین میشود اما در عضله صاف، عملکرد آن ناشناخته است.
- «کالپونین» (Calponin): مولکولهای کالپونین ممکن است به همان اندازه اکتین وجود داشته باشند و پیشنهاد شده که پروتئینهایی باربر هستند.
- «کالدسمون» (Caldesmon): به نظر میرسد که کالدسمون در اتصال اکتین، میوزین و تروپومیوزین نقش دارد، در نتیجه توانایی عضلات صاف را برای حفظ انقباض افزایش میدهد.

دستگاه های انقباضی عضله صاف
میوزین و اکتین بخشهای انقباضی زنجیرههای ساختارهای کششی هستند که هم در میان بافت ماهیچه ای صاف و هم در بین آنها وجود دارند. رشتههای اکتین واحدهای انقباضی به اجسام متراکم متصل میشوند. اجسام متراکم غنی از آلفا اکتینین هستند و به رشتههای بینابینی متصل هستند (که عمدتا از ویمتین و دسمین تشکیل شدهاند)، بنابراین به نظر میرسد که به عنوان لنگرهایی عمل میکنند که رشتههای نازک میتوانند به آنها نیرو وارد کنند.
اجسام متراکم با بتا اکتین که در اسکلت سلولی یافت میشود در ارتباط هستند و این نشان میدهد که اجسام متراکم ممکن است تنشها را در هر دو دستگاه انقباضی و اسکلت سلولی هماهنگ کنند. اجسام متراکم زیر میکروسکوپ الکترونی تیرهتر به نظر میرسند و بنابراین گاهی اوقات به عنوان متراکم الکترونی توصیف میشوند.
رشتههای بینابینی از طریق اجسام متراکم به رشتههای بینابینی دیگر و در نهایت به اتصالات چسبنده غشای سلول عضلانی صاف به نام سارکولما متصل میشوند. اتصالات چسبنده از تعداد زیادی پروتئین شامل آلفا اکتینین، وینکولین و اسید اسکلت سلولی تشکیل شدهاند. اتصالات چسبنده در اطراف باندهای متراکم پراکنده شدهاند که به صورت دنده به دور سلولهای عضله صاف پیچیدهاند.
نوارهای متراکم (یا پلاکهای متراکم) با مناطق غشای حاوی «حفرههای غشایی» (Caveolae) متعدد و متناوب هستند. وقتی کمپلکسهای اکتین و میوزین منقبض میشوند، نیرو از طریق رشتههای بینابینی متصل به این باندهای متراکم، به سارکولما منتقل میشود. در طول انقباض، یک سازماندهی مجدد فضایی دستگاه انقباضی برای بهینهسازی توسعه نیرو وجود دارد. بخشی از این سازماندهی مجدد، متشکل از فسفوریلاسیون ویمیتین در جایگاه آمینواسید سرین ۵۶ توسط یک کیناز فعال شده با p21 است که منجر به جدا شدن پلیمرهای ویمنتین میشود.
در بعضی از انواع بافت ماهیچه ای طول و تعداد رشتههای میوزین و تغییر بین انقباض و آرامش این رشتهها با تغییر نسبت اکتین به میوزین، پویا و متغیر است. سلولهای عضلانی صاف منزوی جدا شده به صورت مارپیچی منقبض میشوند و سلولهای عضلانی صاف قابل انعطافپذیری که به شیشه چسبیدهاند، مناطقی از فعل و انفعالات پروتئین انقباضی را در امتداد محور بلند با انقباض سلول نشان میدهد. بافت ماهیچه ای صاف اغلب نیاز به انقباض دارد بنابراین انعطافپذیری یکی از نیازهای مهم این بافت است.
سلولهای عضله صاف یک ماتریکس خارج سلولی پیچیده حاوی کلاژن (کلاژنهای نوع I و III غالب هستند)، الاستین، گلیکوپروتئینها و پروتئوگلیکانها را ترشح میکنند. عضله صاف یک الاستین و گیرنده کلاژنی ویژه دارد که با پروتئینهای ماتریکس خارج سلولی میانکنش دارند. این فیبرها با ماتریکس خارج سلولی خود در «ویسکوالاستیسیته» (Viscoelasticity) (خاصیتی بین ویسکوزیته و کشسانی) مشارکت دارند. برای مثال عروق خونی بزرگ ویسکوالاستیک هستند، مانند یک «ويندكسل» (Windkessel) عمل میکنند و باعث گسترش انقباض بطن و صاف کردن جریان ضرباندار میشوند و عضله صاف «میانلایه» (Tunica Media) در این خاصیت نقش دارد.
حفرههای غشایی عضلات
سارکولم دارای حفرههای غشایی است که میکرودامینهای رافتهای لیپیدی هستند که برای سیگنالینگ سلولی و کانالهای یونی تخصص یافتهاند. این تهاجمات در سارکوپلاسم حاوی اجزای زیر هستند:
- میزبان گیرنده: پروستاسیکلین، اندوتلین، سروتونین، گیرندههای موسکارینی، گیرندههای آدرنرژیک
- پیامرسانهای ثانویه: آدنیلات سیکلاز، فسفولیپاز C
- G پروتئینها: RhoA و G آلفا
- کینازها: rho kinase-ROCK، پروتئین کیناز C، پروتئین کیناز A
- کانالهای یونی: کانالهای کلسیم نوع L، کانالهای پتاسیم حساس به ATP، کانالهای پتاسیم حساس به کلسیم
حفرهها غالباً به شبکه سارکوپلاسمی یا میتوکندری نزدیک هستند و پیشنهاد میشود که برای سازماندهی مولکولهای سیگنالینگ در غشا عمل میکنند.
انقباض عضله صاف
انقباض عضله صاف با روی هم لغزیدن رشتههای اکتین و میوزین ایجاد میشود. انرژی لازم برای این اتفاق با هیدرولیز ATP فراهم خواهد شد. عملکردهای میوزین به عنوان یک ATPase استفاده از ATP برای تولید یک تغییر کنفورماسیون مولکولی قسمتی از میوزین و ایجاد حرکت است.
حرکت رشتهها روی یکدیگر زمانی اتفاق میافتد که سرهای کروی رشتههای میوزین با رشتههای اکتین متصل شده و میانکنش دهند تا پلهای متقاطعی تشکیل شوند. سر میوزین کج شیب پیدا میکند و به طول کمی دبین ۱۰ تا ۱۲ نانومتر روی رشته اکتین کشیده میشود. سپس سرهای میوزین از اکتین رها میشوند و تغیرر زاویه میدهند تا در یک نقطه دیگر به اندازه ۱۰ تا ۱۲ نانومتر روی اکتین جابجا شوند. میوزینها میتوانند دوباره به مولکول اکتین متصل شوند و به جلو حرکت کنند. این فرآیند «چرخه پل متقاطع» (Crossbridge Cycling) گفته میشود و در تمام عضلات مشابه است.
برخلاف عضله قلبی، عضله صاف فاقد پروتئین متصل به کلیسم تروپونین است و به جای آن انقباض با یک فسفریلاسیون تنظیم شده با کلسیم در میوزین آغاز خواهد شد. چرخه پل متقاطع باعث انقباض کمپلکسهای میوزین و اکتین میشود و به نوبه خود باعث افزایش انقباض در طول کل زنجیره ساختار انقباضی میشود که درن هایت منجر به انقباض کل بافت ماهیچه ای صاف خواهد شد.
انقباض تونیک و فازیک عضله صاف
عضله صاف ممکن است به صورت فیزیکی با انقباض سریع و آرامش یا به صورت تونیک با انقباضات آرام و ثابت منقبض شود. دستگاه تناسلی، گوارشی، تنفسی و ادراری، پوست، چشم و عروق همگی حاوی این نوع عضلات تونیک هستند. این نوع بافت ماهیچه ای صاف میتواند نیرو را برای مدت طولانی و فقط با صرف کمی انرژی حفظ کند. تفاوتهایی در زنجیرههای سبک و سنگین میوزین وجود دارد که با این تفاوت در الگوهای انقباضی و سینتیک انقباض بین عضله صاف تونیک و فازیک ارتباط دارد.
جفت های انقباضی برانگیختگی
یک عضله صاف با محرک خارجی تحریک میشود که به انقباض میانجامد. در ادامه مطلب هر مرحله با جزئیات توضیح داده شدهاند.

محرک های القایی و فاکتورهای انقباض عضله صاف
عضله صاف ممکن است به صورت تصادفی منقبض شود (از طریق کانالهای یونی) یا همانطور که در رودهها سلولهای ضربانساز ویژه وجود دارند، «سلولهای بینابینی کاخال» (Interstitial Cells Of Cajal) انقباضات هماهنگ ایجاد میکنند. همچنین انقباض و آرامش میتوانند توسط تعدادی از عوامل فیزیو شیمیایی ایجاد شوند (به عنوان مثال هورمونها، داروها، انتقال دهندههای عصبی به ویژه از سیستم عصبی خودمختار).
عضلات صاف در مناطق مختلف عروق خونی، مجاری هوایی و ریهها، کلیهها و واژن در بیان کانالهای یونی، گیرندههای هورمونی، مسیرهای سیگنالینگ سلولی و سایر پروتئینهای تعیین کننده عملکرد متفاوتی دارند.
مواد خارجی مؤثر در انقباض عضله صاف
برای مثال عروق خونی پوست، لوله گوارشی، کلیه و مغز به نورواپینفرین و اپینفرین (از تحریک سیستم سمپاتیک یا مدولای آدرنال) باعث انقباض عروقی میشوند (این پاسخ از طریق گیرندههای آدرنرژیک آلفا-۱ هدایت میشود). در هر حال، عروق خونی در عضله اسکلتی و عضله قلبی به این کاتکولآمینها با اتساع عروق پاسخ میدهند، چون دارای گیرندههای بتا آدرنرژیک هستند.
بنابراین تفاوتی در انتشار گیرندههای آدرنرژیک که تفاوت اینکه چرا عروق خونی از مناطق مختلفی به یک ماده نوراپینفرین/اپینفرین به طور متفاوتی پاسخ میدهند و همچنین تفاوتهایی که به دلیل اختلاف در مقدار این کاتکولآمینها که آزاد میشوند و حساسیت گیرندههای مختلف به انقباض بافت ماهیچه ای را توضیح میدهد. به طور کلی، عضلات صاف عروق خونی به کربندی اکسید با اتساع عروق و به اکسیژن با انقباض واکنش میدهند.
عروق خونی ریه منحصر به فرد هستند چون در مقابل فشار اکسیژن متسع و زمانی که اکسیژن خون کاهش مییابد منقبض میشوند. برونشیول، عضله صافی که در مجاری تنفسی ریه قرار دارد، در صورت کمبود دی اکسید کربن متسع و با دی اکسید کربن بالا دچار انقباض عروقی میشود. این واکنشها به کربین دی اکسید توسط عروق خونی ریه و عضلات صاف مجاری برونشیول به سیستم دم و بازدم ریهها کمک میکند. بافتهای عضلانی دیگری که در ریه وجود دارند، شبکههای سارکوپلاسمی اندکی دارند، بنابراین انقباض و انبساط آنها با وابستگی به کلسیم داخل و خارج سلول تفاوت دارد.
انتشار پالس انقباضی
برای ثابت نگاه داشتن ابعاد اندام در مقابل فشار، سلولها با اتصالات چسبنده به یکدیگر چفت شدهاند. در نتیجه، سلولها از نظر فیزیکی به سلولدیگر جفت هستند و انقباض در یک سلول درجاتی از انقباض را به سلولهای اطراف خود منتقل میکند.
اتصالات شکافدار، سلولهای متصل شده را از نظر ترکیبات شیمیایی و الکتریکی درون آنها، به یکدیگر مرتبط میکند به طور مثال با انتقال یون کلسیم یا پتانسیل عمل بین سلولهای ماهیچه صاف را انتقال میدهد. عضله صاف تک واحدی دارای تعداد زیادی اتصالات شکافدار است و ایت بافت اغلب به شکل صفحات یا باندهایی که دستهجمعی منقبض میشوند سازمان یافته است.
رشد و بازسازی عضله صاف
مکانیزمی که در آن عوامل خارجی رشد و بازآرایی را تحریک میکنند هنوز کاملاً درک نشدهاند. تعدادی از فاکتورهای رشد و عوامل نوروهومورال بر رشد و تمایز عضلات صاف تأثیر می گذارند. نشان داده شده است که گیرنده ناچ و مسیر سیگنالینگ سلولی برای تشکیل عروق و رگها ضروری هستند. تکثیر، در پاتوژنز تصلب شرایین نقش دارد و توسط اکسید نیتریک مهار میشود.
عضله صاف به طور معمول منشأ مزودرمی دارد، پس از ایجاد فیبرهای عضلانی و در فرآیندی که به عنوان میوژنز شناخته میشود. با این حال، عضله صاف در شریانهای آئورت و ریوی (شریانهای بزرگ قلب) از اکتومزانشیم تاج عصبی مشتق شده است، اگرچه عضله صاف عروق کرونر منشأ مزودرمی دارد.
عضله صاف در بی مهره ها
در عضله صاف بی مهرهها، انقباض با اتصال مستقیم کلسیم به میوزین و سپس با حرکت سریع از پلهای متقاطع، نیرو ایجاد میکند. مانند مکانیسم عضله صاف مهرهداران، یک مرحله جذب پایین کلسیم و انرژی کم وجود دارد. این فاز پایدار به یک پروتئین گیرنده شباهت دارد که به میوزین زنجیره سبک کیناز و پروتئین الاستیک تیتین شباهت داشته و Twitchin نام دارد. صدف و دیگر نرمتنان دوکاسهای، از این مرحله انقباض ماهیچه صاف استفاده میکنند تا پوسته خود را برای مدت طولانی با مصرف کم انرژی بسته نگه دارند.










سلام تشکر براي من خيلي خوب بود.
سلام لطفا رفرنس مطالب تون رو هم مینویسید؟
سلام، وقت شما بخیر؛
در انتهای کلیه مطالب مجله فرادرس و بعد از بخش معرفی مطالب و آموزشهای مرتبط، منابع هر مطلب به صورت کامل ذکر شدهاند.
از اینکه با مجله فرادرس همراه هستید از شما بسیار سپاسگزاریم.