



پرایمر در ژنتیک چیست؟ – به زبان ساده
پرایمر در ژنتیک یا آغازگر، توالی DNA کوتاه و تک رشتهای است که معمولاً در واکنش زنجیرهای پلیمراز (PCR) استفاده میشود. در روش PCR، از یک جفت آغازگر استفاده میشود تا با DNA نمونه هیبرید شده و ناحیه DNA تقویت شود. از آغازگرها به عنوان الیگونوکلئوتیدها نیز یاد میشود. شما میتوانید از یک آغازگر برای تعیین توالی رشتههای نوکلئوتیدی استفاده کنید، جایی که میخواهید یک منطقه بسیار خاص را هدف قرار داده و سپس تجزیه و تحلیل را در طول آن مولکول DNA انجام دهید.


پرایمر در ژنتیک چیست؟
«آغازگر» (Primer) یا پرایمر در ژنتیک یک توالی کوتاه نوکلئیک اسید است که یک نقطه شروع برای سنتز DNA فراهم میکند. در موجودات زنده، آغازگرها رشتههای کوتاه RNA هستند. قبل از تکثیر DNA، یک آغازگر باید توسط آنزیمی به نام پریماز که نوعی RNA پلیمراز است، سنتز شود. سنتز یک آغازگر ضروری است زیرا آنزیمهایی که DNA را سنتز میکنند و DNA پلیمرازها نامیده میشوند، فقط میتوانند نوکلئوتیدهای DNA جدید را به یک رشته نوکلئوتید موجود متصل کنند. آغازگر به مجموعه کوچکی از نوکلئوتیدهای DNA اشاره دارد که طول آنها معمولاً 18 تا 24 نوکلئوتید است و از آن میتوان برای بسیاری از فرایندهای تکثیر آزمایشگاهی استفاده کرد.
آغازگرها قبل از تکمیل تکثیر DNA برداشته میشوند و شکافهای توالی DNA توسط DNA پلیمرازها پر میشود. در آزمایشگاه، دانشمندان میتوانند آغازگرهای DNA با توالیهای خاص را که به توالیهای یک مولکول DNA تکرشته متصل میشوند، طراحی و سنتز کنند. این آغازگرهای DNA معمولاً برای انجام واکنش زنجیرهای پلیمراز برای کپی کردن قطعات DNA یا برای تعیین توالی DNA استفاده میشوند. طراحی آغازگر یک گام اساسی در کاربرد فناوریهای مبتنی بر انواع PCR در بیان ژن و تجزیه و تحلیل تنوع ژنتیکی است. پروفایل mRNA برای شناسایی ژنهای جدید، تعیین عملکرد ژنها و روشن کردن شبکههای ژنتیکی بسیار مهم است. نمایش افتراقی، چند شکلی طول قطعه تقویت شده cDNA که به صورت (cDNA - AFLP) نشان داده میشود و تکنولوژی «ریزآرایه» (Microarray) به طور گستردهای برای پروفایل mRNA های گیاهی استفاده شده است.

نمایهسازی چند شکلی DNA برای نقشه برداری ژن، انتخاب گیاهان زراعی با کمک مارکر و مطالعات تنوع مولکولی ضروری است. تکنیکهای مبتنی بر PCR مانند AFLP و «ریزماهوارهها» (Microsatellites) یا تکرار توالی ساده (SSR) نیز نقش مهمی در پروفایل DNA گیاه داشتهاند. پرایمر در ژنتیک از اجزای اساسی سیستمهای مبتنی بر PCR و همچنین سیستمهای ریزآرایه مدرن است که از «کاوشگرهای» (Probes) مناسب که با تکثیر PCR بدست میآیند استفاده میکند.
انواع پرایمرهای DNA و RNA
موجودات زنده فقط از آغازگرهای RNA استفاده میکنند، در حالی که آغازگرهای مورد استفاده در آزمایشگاه معمولاً آغازگرهای DNA هستند. دانشمندان به دلایل مختلف یا متنوعی از پرایمرهای DNA به جای آغازگرهای RNA استفاده میکنند. آغازگرهای DNA به مراتب پایدارتر بوده و راحتتر ذخیره میشوند و برای شروع سنتز به آنزیمهای رایجتری احتیاج دارند. اتصال آغازگرهای DNA یا RNA به رشته الگو، آنزیم مسئول سنتز DNA را فعال میکند، DNA پلیمراز با شروع افزودن نوکلئوتیدها به انتهای ’۳ - هیدروکسیل (انتهای ’۳) یک اسید نوکلئیک موجود بر روی پرایمر، باعث طویل شدن و تکثیر رشته اصلی میشود. در جدول زیر ویژگیهای مختف پرایمرهای DNA و RNA را با هم مقایسه کردهایم.
| ویژگیها | آغازگرهای DNA | آغازگرهای RNA |
| استفاده | در شرایط آزمایشگاهی: تکثیر PCR، تعیین توالی DNA، کلونینگ و موارد دیگر | در بدن موجودات زنده: همانند سازی DNA |
| واکنش | تکثیر وابسته به دما است و به پروتئین کمتری نیاز دارد. | تکثیر یک واکنش کاتالیزوری وابسته به آنزیم است که به چندین پروتئین نیاز دارد. |
| طول | 18 تا 24 نوکلئوتید | 10 تا 20 نوکلئوتید |
| نحوه ایجاد | توسط پژوهشگران سنتز شیمیایی شده است. | توسط پرایماز (نوعی RNA پلیمراز) در بدن موجود زنده ساخته شده است. |
| قابلیت زیستن | عمر طولانیتر، با ثباتتر | عمر کوتاهتر، واکنش پذیرتر |
پرایمر در همانندسازی DNA
این نقشی است که آغازگرهای RNA در سنتز DNA ایفا میکنند و اهمیت پرایمر در ژنتیک را بسیار بالا میبرند. در بدن موجودات زنده (In vivo) این همانندسازی DNA یا سنتز ژن است که سنگ بنای وراثت را تعیین میکند و در آن مواد ژنتیکی که از سلولهای اصلی به ارث رسیدهاند به منظور رشد، تقسیم و تمایز سلول کپی میشوند. تکثیر DNA از مبدا همانندسازی آغاز شده که این مبدا توسط پروتئینهایی به نام آغازگرها شناسایی میشوند که روی DNA قفل شده و دهانههای کوچکی در «مارپیچ DNA دو رشتهای» (dsDNA) ایجاد میکنند. در این شکستها، آنزیمهای هلیکاز پیوندهای بین رشتههای دوتایی را میشکند و DNA تک رشتهای (ssDNA) را در انتهای Y شکل به نام «چنگال تکثیر DNA» یا DNA replication fork قرار میدهند.
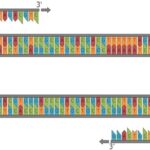
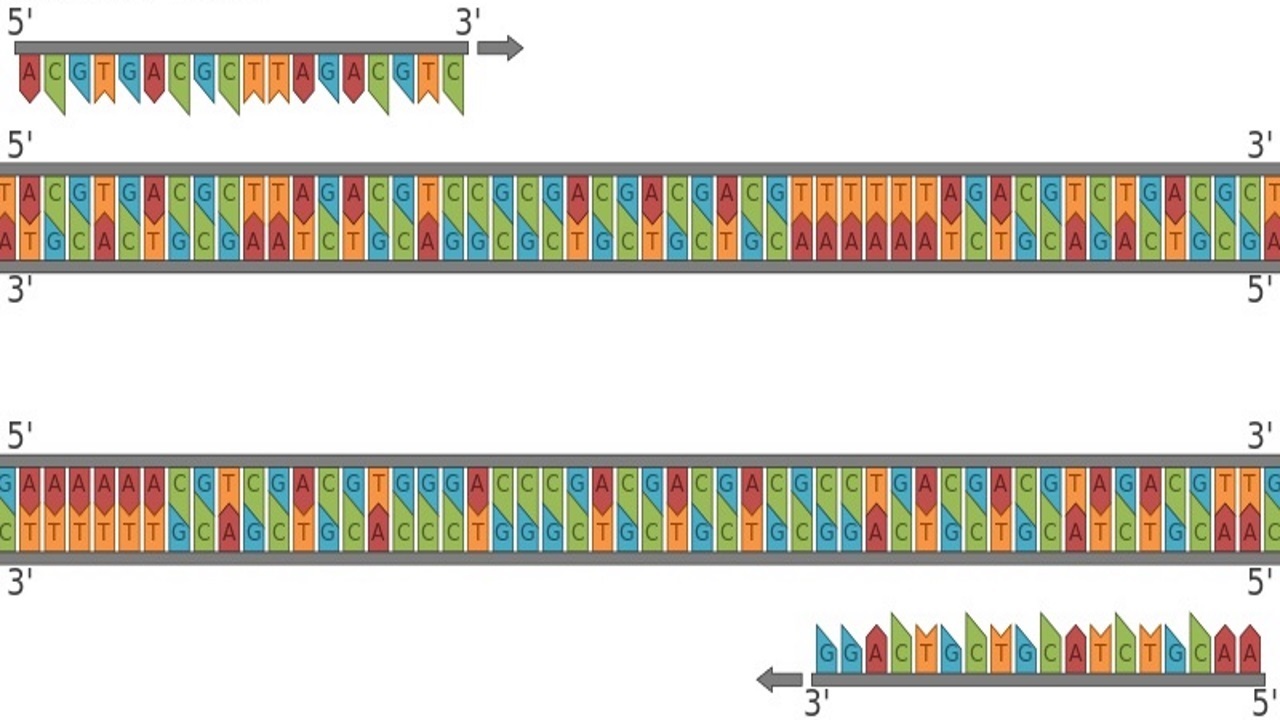
چنگال همانندسازی همان جایی است که تکثیر DNA واقعاً رخ خواهد داد، و دو رشته DNA والدی را در معرض قرار میگیرند تا به عنوان الگوی DNA عمل کنند (رشتههای خاکستری تصویر زیر). در مرحله بعدی، آنزیم پرایماز (RNA primase) آغازگر RNA را روی هر یک از رشتههای الگوی اصلی قرار میدهد، و انتهای ’3 را برای شروع تشکیل یک رشته پلی نوکلئوتیدی جدید فراهم میکند. سپس DNA پلیمراز متصل شده و با ایجاد رشتههای دختر (رشتههای بنفش) سنتز آغاز میشود. در این تصویر، رشته دختر بالایی به عنوان رشته پیشتاز خوانده میشود و در جهت ’5 به ’۳ یعنی از راست به چپ در معرض همانندسازی قرار میگیرد و اجازه میدهد تا سنتز DNA به طور مداوم هنگام باز شدن مارپیچ رخ دهد. سنتز رشته پیرو در رشته دختری پایین در جهت ’5 به ’3 چپ به راست اتفاق میافتد، جایی که سنتز DNA باید با باز شدن مارپیچ به طور مداوم شروع شود.
این باعث ایجاد بسیاری از قطعات کوتاه از محصولات سنتز شده میشود که به عنوان قطعات اوکازاکی شناخته میشوند و طول آنها به طور متوسط 150 تا 200 نوکلئوتید است. هر قطعه اوکازاکی حاوی یک اتصال RNA - DNA به عنوان یک نتیجه از آغازگر RNA است، کشفی که از شواهد مهمی برای تعیین نقش آغازگرهای RNA در همانندسازی DNA بود. قطعات اوكازاكی بايد توسط آنزيم ليگاز به هم متصل شوند كه این آنزیم تشكيل پيوندهای فسفو دی استر كوالانسی بين دو رشته DNA را كاتاليز كرده و آنها را بهم پيوند میدهد.

پرایمرهای Forward و Reverse
آغازگرها فقط به DNA تک رشته متصل میشوند. در تکثیر DNA در داخل بدن، این مورد در چنگال همانند سازی در دسترس قرار میگیرد. در شرایط آزمایشگاهی، دانشمندان میتوانند با دناتوره كردن دی ان ای دو رشتهای با حرارت، دی ان ای تک رشتهای ایجاد كرده و به طور مؤثر پیوندهای هیدروژنی كه دو رشته را به هم میچسباند، از بین ببرند. این دو رشته الگوی خطی را ایجاد میکند که در آن سنتز DNA میتواند به طور مداوم روی هر دو رشته رخ دهد که اغلب به جای اصطلاح رشتههای پیشرو و عقبی، رشتههای بالا و پایین نامیده میشوند. بنابراین یک جفت آغازگر باید استفاده شود، یک پرایمر برای رشته بالا و دیگری برای پایین. جفت آغازگر در انتهای مخالف دنباله متصل میشود، در حالی که انتهای ’3 آنها به سمت یکدیگر است.
آغازگرهای الیگونوکلئوتیدی که در حال طراحی هستند، لازم نیست که کاملاً به رشته الگو پیوند خورند. با این حال ضروری است که انتهای ’3 به طور کامل با DNA الگو مطابقت داشته باشد تا طویل شدن اتفاق بیفتد. پرایمر فوروارد به رشته پایینی که در حال باز شدن از ’۳ به ’۵ است متصل میشود و پرایمر معکوس به رشته بالایی DNA مکمل که از ’5 تا ’3 در حال باز شدن است، متصل میشود. پس از طویل شدن، این واکنش منجر به ساخت دو رشته DNA دو رشتهای جدید میشود که یکی ساخته شده از پرایمر فوروارد و دیگری از آغازگر معکوس ساخته شده است. برای اینکه آغازگرها به رشته الگو متصل شوند، مهم است که مکمل یکدیگر نباشند. آغازگرهای مکمل یکدیگر میتوانند منجر به ایجاد پرایمر - دایمر شوند که دو آغازگر هستند و به جای رشته الگو به یکدیگر متصل میشوند.

طراحی پرایمر در ژنتیک
درک خصوصیات آغازگر برای طراحی آن بسیار مهم است. جنبههای اصلی خصوصیات یک پرایمر در ژنتیک شامل اختصاصیت، دمای ذوب (Tm) و همسانی درون پرایمری یا بین آن است. اختصاصیت پرایمر بیشتر توسط توالی های ’۳ آنها تعیین میشود. عدم تطابق تک نوکلئوتید داخل پرایمر تأثیر معنی داری بر عملکرد محصول PCR ندارد در حالی که عدم تطابق انتهای ’3 یک پرایمر، به ویژه عدم تطابق A، G و C، به طور قابل توجهی عملکرد کلی محصول PCR را کاهش میدهد. اختصاصیت اتصال آغازگرها را در واکنشهای PCR تحت دماهای مختلف (با توجه به دمای Tm پرایمر) میتوان ارزیابی کرد و دقت تکثیر DNA را افزایش داد.
بنابراین گنجاندن 8 تا 10 باز منحصر به فرد در انتهای ’۳ آغازگر بسیار مهم است. اختصاصیت جایگاه اتصال آغازگر را میتوان با انجام یک جستجوی همسانی توالی (به عنوان مثال Blastn) از طریق تمام توالیهای الگوی شناخته شده در پایگاه داده ژنوم عمومی مانند مرکز ملی اطلاعات بیوتکنولوژی (NCBI) بررسی کرد. برای اطمینان از اتصال خاص آغازگر به الگوی DNA، پرهیز از حضور 4 عدد یا بیشتر G (گوانین) یا C (سیتوزین) در یک ردیف در انتهای ’3 نیز مهم است. Tm یک پرایمر در ژنتیک با طول آغازگر، محتوای GC و ترکیب نوکلئوتید تعیین میشود و در صورت زیاد بودن تعداد GC ها مشکلاتی را در روند تکثیر به دنبال خواهد داشت.
نکات مهم در طراحی پرایمر
واکنش زنجیرهای پلیمراز (PCR) به عنوان یکی از مهمترین اختراعات قرن بیستم در زیست شناسی مولکولی به طور گستردهای انجام میشود. امروزه با تکثیر مقادیر کمی از مواد ژنتیکی از طریق PCR میتوان کارهایی مانند شناسایی و دستکاری DNA، شناسایی ارگانیسمهای عفونی (از جمله ویروسهایی که باعث ایدز، هپاتیت و سل میشوند)، تغییرات ژنتیکی از جمله جهش در ژنهای انسانی و سایر کارهای مختلف را انجام داد. PCR شامل سه مرحله است: «دناتوراسیون» (Denaturation)، «اتصال» (Annealing) و «طویل شدن» (Extension).

به این ترتیب که ابتدا ماده ژنتیکی دناتوره میشود و مولکول های DNA دو رشتهای را به تک رشتههای DNA تبدیل میکند. آغازگرها سپس به مناطق مکمل مولکولهای تک رشتهای متصل میشوند. در مرحله سوم، آنها با عمل DNA پلیمراز گسترش مییابند. تمام این مراحل به دما حساس هستند و انتخاب معمول دما به ترتیب 94 درجه سانتیگراد، 60 درجه سانتیگراد و 7۲ درجه سانتیگراد است. طراحی خوب آغازگر برای واکنشهای موفق ضروری است، ملاحظات مهم طراحی پرایمر در ژنتیک که در زیر توضیح داده شده، کلیدی برای تکثیر اختصاصی با عملکرد بالا هستند.
طول پرایمر
در طراحی پرایمر در ژنتیک به طور کلی پذیرفته شده است که طول بهینه آغازگرهای پی سی آر 18 تا 22 باز است. این طول برای اختصاصیت مناسب بوده و همچنین به اندازه کافی کوتاه است تا پرایمرها بتوانند به راحتی در دمای اتصال به رشته الگوی DNA متصل شوند. باید توجه شود که طول پرایمر خیلی کوتاه نباشد زیرا آغازگرهای کوتاه میتوانند منجر به اتصال غیر اختصاصی و در نتیجه محصولات PCR نادرست شوند. همچنین نباید طول آنها بیش از ۳۰ نوکلئوتید باشد زیرا ممکن است سرعت ترکیب شدن را کاهش دهند. طول و ترکیب پرایمر در ژنتیک مستقیماً بر دمای ذوب و اتصال پرایمر تأثیر میگذارد.
دمای ذوب پرایمر
دمای ذوب (Tm) پرایمر در ژنتیک بر اساس تعریف دمایی است که در آن نیمی از دو رشته DNA از هم فاصله گرفته تا به صورت تک رشته درآیند و این دما پایداری دوبلکس DNA را نشان میدهد. پرایمرهای با دمای ذوب محدوده ۵۰ تا ۶۰ درجه سانتیگراد با حداکثر ۵ درجه اختلاف دما با هم به طور کلی بهترین نتیجه را دارند. اگر دمای ذوب هر آغازگر نسبت به دمای اتصال برای واکنش بسیار زیاد یا کم باشد، هیبریداسیون اشتباه ممکن است رخ دهد. آغازگرهای با دمای ذوب بالای 65 درجه سانتیگراد تمایل به ایجاد اتصال مجدد و ایجاد ساختارهای ثانویه دارند. محتوای GC توالی رشته DNA، به درستی میتواند نشانگر دمای ذوب پرایمر (Tm) باشد. برای محاسبه دمای ذوب پرایمر به صورت کلی برای پرایمرهای با طول ۲۰ نوکلئوتید و کمتر از فرمول زیر استفاده میشود.
Tm = 4 (G+C) + 2(A+T)
آغازگرهای با محتوای G - C بالاتر دمای ذوب بیشتری از خود نشان میدهند زیرا شکستن پیوندهای هیدروژنی G - C دشوارتر از پیوندهای هیدروژنی A - T است، بنابراین برای ذوب شدن به انرژی بیشتری (دمای بالاتر) نیاز است. برای پرایمرهای با اندازه بلندتر با طول و تعداد نوکلئوتید بیشتر فرمول بالا کاربرد نداشته و فرمول محاسبه دمای ذوب یا Tm پرایمر از لحاظ ترمودینامیکی از «تئوری ترمودینامیک نزدیکترین همسایه» (Nearest Neighbor Thermodynamic Theory) استفاده میشود که توسط نرم افزارهای تخصصی قابل محاسبه است. فرمولهای مربوط به این تئوری به صورت خلاصه در ادامه بیان شدهاند.
فرمول محاسبه دمای ذوب پرایمر برای پرایمرهای با طول بیش از ۲۰ نوکلئوتید:
Tm (K)= ΔΗ/ΔS + R ln (C)
Tm (°C)= ΔΗ/ΔS + R ln (C) - 273/15
در عبارت ΔH (kcal / mole)، باید گفت که H آنتالپی است. آنتالپی مقدار انرژی گرمایی است که توسط مواد به وجود میآید. ΔH تغییر در آنتالپی است، در فرمول فوق، ΔH با جمع کردن مقادیر آنتالپی تمام جفت دی - نوکلئوتیدهای نزدیکترین همسایه هر نوکلئوتید بدست میآید و C مقدار غلظت کل نوکلئوتیدهای موجود است.
در عبارت ΔS (kcal / mole)، باید گفت که S میزان اختلالی است که سیستم نشان میدهد و آنتروپی نامیده میشود. در اینجا با جمع کردن مقادیر آنتروپی تمام جفت دی - نوکلئوتیدهای نزدیکترین همسایه هر نوکلئوتید بدست میآید. با توجه به اینکه پارامترهای نزدیکترین همسایه از مطالعات ذوب DNA به دست آمد، یک اصلاح اضافی نمک اضافه میشود.
ΔS (salt correction) = ΔS (1M NaCl ) + 0/368 × N × ln ([Na+])
که در آن N تعداد جفتهای نوکلئوتید در آغازگر است. (طول پرایمر منهای یک).
[Na +] معادل نمک در میلی مولار است. یونهای دو ظرفیتی مانند منیزیم دارای ۲ بار مثبت در مقایسه با یونهای یک ظرفیتی مانند Na + تأثیر مهمی بر ترکیبی DNA و RNA دارند. در نتیجه منیزیم به منظور بهبود و تسریع چین خوردن DNA به راحتی در بسیاری از انواع حالات چین خوردگیهای رشته DNA استفاده میشود.
محاسبه [Na +]:
[Na +] = (غلظت یون تک ظرفیتی) + ۴ × Mg2+ آزاد
دمای اتصال پرایمر
هنگام تصمیم گیری در مورد «دمای اتصال» (Ta) باید مستقیماً Tm را در نظر بگیرید. دمای ذوب پرایمر برآورد ثبات ترکیبی DNA - DNA بوده و برای تعیین دمای اتصال بسیار مهم است. Ta خیلی بالا ناکافی بودن هیبریداسیون الگو - پرایمر و در نتیجه بازده کمتر محصول PCR را موجب میشود. Ta خیلی پایین ممکن است منجر به محصولات غیر اختصاصی ناشی از تعداد زیاد عدم تطابق نوکلئوتید شود. تحمل عدم تطابق نوکلئوتیدی بیشترین تأثیر را در اختصاصیت PCR دارد.
Ta = 0/3 × Tm (پرایمر) + 0/7 Tm (محصول) - 14/9
که در این فرمول Tm (پرایمر) = دمای ذوب پرایمرها بوده و Tm (محصول) = دمای ذوب محصول PCR است. دمای اتصال بهینه به صورت تجربی کشف میشود، اما به طور کلی حدود ۵ تا ۱۰ درجه سانتیگراد کمتر از دمای ذوب آغازگر (Tm) است. اگر دمای اتصال پرایمر خیلی زیاد باشد، احتمالاً پرایمرها به اندازه کافی متصل نخواهند شد و در نتیجه آمپلیکونها به مقدار کم یا صفر تکثیر میشوند. اگر دمای اتصال آغازگر خیلی کم باشد، پرایمرها ممکن است شروع به تشکیل ساختارهای سنجاق سری یا اتصال به مناطق خارج از توالی هدف کنند که منجر به تولید محصولات غیر اختصاصی و غیر دقیق در PCR میشود. یکی از روشهای معمول برای یافتن دمای بهینه اتصال اولیه، انجام یک PCR با گرادیان دمایی یا PCR با گرادیان حرارتی است که از حدود 5 درجه سانتیگراد کمتر از پایینترین دمای ذوب جفت پرایمر شروع میشود.

محتوای GC چه نقشی در طراحی پرایمر دارد؟
همانطور که میدانیم بین نوکلئوتیدهای گوانین و سیتوزین پیوندهای هیدروژنی سهگانه برقرار شده در حالی که بین بازهای آدنین و تیمین پیوند هیدروژنی دوگانه برقرار میشود. وجود تناسب کافی بین بازهای گفته شده در طراحی پرایمر جزو نکات بسیار مهم آن است. چراکه در صورت زیاد بودن محتوای GC دمای اتصال بالا رفته و در روند تکثیر DNA اختلال ایجاد میشود. بنابراین محتوای GC (تعداد نوکلئوتیدهای G و C در پرایمر به عنوان درصدی از تعداد کل بازهای قطعه DNA مورد نظر) پرایمر باید 40 تا 60 درصد باشد. در 5 باز آخر در انتهای ’۳ پرایمر، باید حداقل 2 باز G یا C وجود داشته باشد که این به عنوان «گیره GC» یا GC Clamp شناخته میشود.
ساختارهای ثانویه پرایمر
وجود ساختارهای ثانویه پرایمر تولید شده توسط فعل و انفعالات بین مولکولی یا درون مولکولی میتواند منجر به کم شدن یا عدم بازده تولید محصول واکنش PCR شود. آنها اثر منفی بر روی اتصال رشته الگوی متصل شده به پرایمر و در نتیجه تکثیر DNA هدف دارند. این ساختارها در دسترس بودن آغازگرهای واکنش را تا حد زیادی کاهش میدهند. در ادامه انواع ساختارهای ثانویهای که باعث اختلال در روند اتصال پرایمر به رشته الگو میشوند را بررسی میکنیم.
ساختار سنجاق سری چیست؟
این ساختار ثانویه در اثر فعل و انفعالات درون مولکولی داخل پرایمر تشکیل میشود و باید از آن اجتناب شود. اغلب این نوع سختار در صورت وجود همسانی درون آغازگر ایجاد شده و در آن پرایمر منفرد به خودی خود جمع میشود، یا اگر دمای ذوب آغازگر کمتر از دمای اتصال واکنش باشد این حالت رخ میدهد. ساختارهای سنجاق سری انتهای ’3 از نامطلوبترین آنها هستند زیرا پرایمر کاملاً روی خود جمع شده است، در این حالت ۳ باز اول و آخر به هم متصل میشوند. در حالت مطلوب یک ساختار سنجاق سری انتهایی ’3 با ΔG منفی ۲ کیلو کالری بر مول و یک ساختار سنجاق سری داخلی با ΔG منفی ۳ کیلو کالری بر مول به طور کلی قابل تحمل است. اگر دلتا جی بیش از این منفی باشد ممکن است پرایمر در واکنش PCR از هم باز نشود.

تعریف ΔG
انرژی آزاد گیبس (G) مقیاس میزان کاری است که میتواند از یک فرآیند با فشار ثابت استخراج شود. این مقیاس اندازهگیری خود به خودی بودن واکنش است. ثبات ساختار سنجاق سری معمولاً با مقدار ΔG آن، انرژی مورد نیاز برای شکستن ساختار ثانویه، نشان داده می شود. مقدار منفی بیشتر برای ΔG نشانگر ساختارهای سنجاق سری مستحکم و نامطلوب است. وجود این ساختارهای سنجاق سری در انتهای ’3 بر روی واکنش تأثیر منفی میگذارد.
ΔG = ΔH – TΔS
ساختار خود - دایمر
یک «پرایمر - خود - دایمر» (Primer - Self - Dimer) در اثر فعل و انفعالات بین مولکولی بین دو پرایمر همسان در ژنتیک ایجاد میشود، جایی که آغازگر با خودش همولوگ است. در واقع هنگامی که پرایمرها به جای رشته الگو با یکدیگر اتصال برقرار میکنند، با همولوژی بین آغازگرها ایجاد میشوند. به طور کلی مقدار بیشتری از پرایمرها در مقایسه با مقدار ژن هدف در PCR استفاده میشود. وقتی آغازگرها دایمرهای بین مولکولی را خیلی راحتتر از متصل شدن به DNA هدف تشکیل میدهند، بازده تکثیر محصول کاهش مییابد. در حالت مطلوب، یک دایمر انتهایی ’3 با ΔG منفی ۵ (۵-) کیلو کالری بر مول و یک خود دایمر داخلی با ΔG منفی ۶ (۶-) کیلو کالری بر مول به طور کلی تحمل میشود.
ساختار دایمر متقاطع
دایمرهای متقاطع پرایمرها با فعل و انفعالات بین مولکولی بین آغازگرهای فوروارد و ریورس، در جایی که همولوگ هستند، تشکیل میشوند. در حالت مطلوب یک دایمر متقاطع انتهایی ’3 با ΔG منفی ۵ (۵-) کیلو کالری بر مول و یک دایمر متقاطع داخلی با ΔG منفی ۶ (۶-) کیلو کالری بر مول به طور کلی تحمل میشود.

تکرارها در توالی
تکرار، یک دی - نوکلئوتید است که بارها و بارها اتفاق میافتد و باید از آن اجتناب شود، زیرا ممکن است باعث اختلال در روند تکثیر شود. به عنوان مثال: ATATATAT. حداکثر تعداد تکرارهای دی نوکلئوتیدی قابل قبول در یک رشته اولیگونوکلئوتیدی 4 دی نوکلئوتید است. همچنین در مورد انتهای ’۳ باید در نظر داشت که برای پایداری حداکثر مقدار ΔG پنج باز از انتهای ’3 است. انتهای ’3 ناپایدار (ΔG منفی کمتر) باعث ایجاد شدن کمتر اتصال کاذب پرایمر میشود.
ساختارهای ثانویه در رشته الگوی DNA
توالیهای اسید نوکلئیک تک رشتهای بسیار ناپایدار بوده و به صورت ترکیبات (ساختارهای ثانویه) جمع میشوند. پایداری این ساختارهای ثانویه رشته الگوی DNA تا حد زیادی به انرژی آزاد و دمای ذوب آنها (Tm) وابسته است. در نظر گرفتن ساختارهای ثانویه DNA الگو در طراحی آغازگرها، به ویژه در qPCR مهم است. اگر پرایمرها بر روی ساختارهای ثانویه و حتی در دمای اتصال پایدار طراحی شوند، نمیتوانند به الگو متصل شوند و بازده تکثیر محصول PCR به طور قابل توجهی تحت تأثیر قرار میگیرد. از این رو، طراحی پرایمرها در مناطقی از الگوهای DNA که در طی واکنش PCR ساختار ثانویه پایدار ایجاد نمیکنند، مهم است.
همسانی متقاطع رشته الگو
برای بهبود اختصاصیت پرایمر در ژنتیک لازم است از به کار بردن مناطق همسانی رشته الگوی DNA اجتناب شود. پرایمرهای ساخته شده برای یک توالی الیگونوکلئوتیدی نباید سایر ژنها را در مخلوط تکثیر کنند. به طور معمول، پرایمر در ژنتیک برای آزمایش اختصاصیت طراحی و سپس برای بررسی اختصاصیت آنها در وب سایت NCBI بلاست میشوند. شما میتوانید الگوهای DNA مورد نظر را در این پایگاه داده BLAST کرده و نرم افزار آن نتایج را تفسیر میکند. با این کار مناطق قابل توجهی را در هر الگو شناسایی کرده و از بروز آنها در هنگام جستجو برای محل مناسب اتصال پرایمر جلوگیری میکند.
بلاست در NCBI
NCBI Blast مجموعه برنامههایی است که در سال 1991 توسط مرکز ملی اطلاعات بیوتکنولوژی ایالات متحده (NCBI) آغاز شده است. «ابزار جستجوی هم ترازی محلی» (BLAST) میتواند مناطق مشابهت بین توالیهای آزمایشی مورد نظر و موجود در پایگاه دادهای را شناسایی کند. با مقایسه توالیهای نوکلئوتیدی با پایگاههای توالی مختلف، این ابزار قادر است گزارشات معنی دار آماری دقیقی را از هر مورد مطابقت یافته ارائه دهد. الگوریتمهای BLAST مختلفی وجود دارد، از جمله مواردی که به شناسایی ژنومها (توالیهای نوکلئوتیدی RNA یا DNA)، بخشهای هدفمند مانند SNP ها و حتی پروتئینها کمک میکند. هر پرسشی که دانشمندان با استفاده از BLAST انجام میدهد برای همیشه ذخیره میشود و یک کتابخانه همیشه در حال رشد از توالیهای مرجع ایجاد میکند که به دقت این ابزار میافزاید.

طراحی پرایمر برای کلونینگ
شبیه سازی مولکولی یک تکنیک مهم است که به دانشمندان امکان میدهد بخشهای خاصی از توالی هدف DNA را در سلولهای میزبان تکثیر کنند. ژن یا قطعه مورد نظر را میتوان در یک وکتور یا حامل کلونینگ قرار داد، به طور کلی یک ژنوم پلاسمید پروکاریوتی دایرهای (که گاهی اوقات وکتور پلاسمیدی نامیده میشود)، میتواند توسط یک سلول گیرنده خاص، معمولاً یک سلول باکتریایی، جذب شود. سپس وکتور از طریق ترکیب مجدد DNA در ژنوم میزبان قرار میگیرد، و همانطور که سلول به طور طبیعی تحت همانندسازی قرار میگیرد، ژن مورد نظر نیز تکثیر میشود یا در واقع کلون میشود.

اما ابتدا پژوهشگران باید ژن هدف خاص را برای کلون سازی جدا کنند. آنها میتوانند این کار را با استفاده از PCR انجام دهند تا بعد از جدا شدن ژن هدف، این ژن را تکثیر کنند.
متناوباً، اگر توالی مورد نظر برای تکثیر در یک پلاسمید یا در توالی دیگری باشد، آنها میتوانند مکانهای محدود کننده توالی را پیدا کنند تا بتوانند آن را با استفاده از آنزیمهای محدود کننده برش دهند، یا بهتر بگوییم توالی را هضم کنند. هر دو روش به دانشمندان اجازه میدهد تا بخش مشخصی از DNA را تولید كنند و سپس میتوانند به صورت ناقل پلاسمید جمع شوند تا برای کلونینگ به ردههای سلولی تبدیل شوند.
طراحی انتهای ’5 پرایمر
علاوه بر مواردی مانند طول پرایمر در ژنتیک که معمولاً طول بین 18 تا 30 نوکلئوتید برای اکثر کاربردهای PCR بهینه است. آغازگرهای کوتاهتر میتوانند منجر به تکثیر محصولات غیر اختصاصی PCR شوند. همچنین باید توجه داشت که در انتهای ’۳ پرایمر لازم است در مواردی مانند تعداد ۳ یا بیشتر نوکلئوتید G یا C وجود داشته باشد تا اتصالات غیر اختصاصی را خنثی کنند، چراکه یک انتهای ’۳ تیمیدین احتمال اتصال غیر اختصاصی پرایمر در ژنتیک را بالا میبرد. به علاوه جفت پرایمرها باید از نظر مکمل بودن با هم در انتهای ’۳ بررسی شوند. این اغلب منجر به تشکیل پرایمر - دایمر میشود. بازهای انتهای ’۵ برای اتصال آغازگر از اهمیت کمتری برخوردار هستند. بنابراین، میتوان عناصر توالی مانند مکانهای اتصال آنزیم محدود کننده را به انتهای ’۵ مولکول پرایمر اضافه کرد.
همانطور که بیان شد اختصاصیت PCR به شدت به دمای ذوب (Tm) پرایمر در ژنتیک بستگی دارد (دمایی که نیمی از آغازگر رشته الگوی DNA متصل شده است). نتایج خوب معمولاً زمانی بدست میآید که Tm برای هر دو آغازگر مشابه باشد ( با ۲ تا ۴ درجه سانتیگراد اختلاف) و بالاتر از 60 درجه سانتیگراد باشد. TM را به صورت سادهتر با استفاده از فرمول زیر میتوان تخمین زد:
Tm = 2°C × (A + T) + 4°C × (C + G)
انتهای ’۵ پرایمر در ژنتیک با انتهای ’۵ ژن مورد علاقه همپوشانی دارد و باید حاوی عناصر زیر باشد:
- جایگاه اتصال آنزیم محدودکننده. توالی آنزیم محدودکننده باید در پرایمر و در وکتور مورد نظر یکسان باشد در واقع با توجه به جایگاههای آنزیمهای محدود کننده موجود در وکتور باید توالی مربوط به آنزیم در پرایمر در نظر گرفته شده و در طراحی پرایمر در ژنتیک اعمال شود. این شرایط در مورد آنزیمهایی که انتهای چسبنده ایجاد میکنند وجود دارد. در مورد آنزیمهای ایجاد کننده انتهای صاف میتوان هر آنزیمی را انتخاب کرد. غالباً آنزیم (Nco I) با توالی اتصال (CCATGG) یا (Nde I) با توالی اتصال (CATATG) انتخاب میشوند زیرا از ATG موجود در این جایگاهها می توان مستقیماً برای ایجاد کدون شروع ATG و یا کدن ATG برای متیونین N - ترمینال استفاده کرد.
- اضافه کردن ’۵ به جایگاه محدودکننده. آنزیمهای محدود کننده داخل DNA را با تاثیرگذاری خیلی کمتری نسبت به انتهای یک رشته برش میدهند. یک انتهای ’۵ در جایگاه اتصال آنزیم محدودکننده با 2 تا 10 نوکلئوتید کارآیی هضم آنزیمی را بسیار افزایش میدهد.
- کدون آغاز. کدون شروع (معمولاً ATG) باید در هنگام طراحی پرایمر درج شود که ژن مورد نظر با برچسب N-terminal یا کاندید همجوشی هنگامی که هنوز یک متیونین N ترمینال وجود دارد بیان نشود. باید بررسی شود که کدون شروع و ژن مورد نظر با یک برچسب نهایی ترمینال N و یا کاندید همجوشی در چارچوب خوانش قرار دارند.
- همپوشانی با ژن مورد نظر. همپوشانی بین آغازگر و ژن مورد نظر باید آنقدر طولانی باشد که Tm حدود ۶۰ درجه سانتیگراد یا بیشتر بدست آورد.

طراحی انتهای ’۳ پرایمر
انتهای ’۳ پرایمر در ژنتیک با انتهای ’۳ رشته DNA مکمل ژن مورد نظر همپوشانی دارد و این انتها نیز باید حاوی جایگاه آنزیم محدودکننده بوده و با جایگاه مورد نظر در داخل وکتور همخوانی داشته باشد. در اینجا نیز شما میتوانید هر آنزیم محدود کنندهای را که پس از برش، انتهای صاف دارد، انتخاب کنید. در این انتها نیز قرار دادن یک انتهای ’۵ در جایگاه اتصال آنزیم محدودکننده با 2 تا 10 نوکلئوتید کارآیی هضم آنزیمی را بسیار افزایش میدهد. نکات مهم دیگر نیز در ادامه بررسی شدهاند:
- کدون پایان. در صورت عدم استفاده از برچسب C-terminal، باید یک کدون توقف (ترجیحاً TAA زیرا نسبت به TAG و TGA کمتر در معرض خواندن توسط آنزیم قرار دارد).
- همپوشانی با رشته مکمل انتهای ’۳ ژن مورد نظر. همپوشانی بین پرایمر در ژنتیک و رشته مکمل انتهای ’۳ ژن مورد نظر باید به اندازه کافی طولانی باشد تا یک Tm 60 درجه سانتیگراد یا بیشتر ایجاد کند. همچنین باید بررسی شود که ژن مورد نظر در یک چارچوب با برچسب C-terminal باشد.
ابزارهای طراحی پرایمر در ژنتیک
در حالت ایدهآل، آغازگر دارای یک Tm در محدوده 50 - 65 درجه سانتیگراد، ترکیب نوکلئوتیدی تصادفی، 40 تا 60 درصد محتوای GC و 18 تا 30 عدد باز طول دارد. برای جلوگیری از ایجاد ساختارهای سنجاق سری (بیش از 3 نوکلئوتید مکمل در داخل پرایمر) یا ساختارهای پرایمر - دایمر (بیش از ۳ نوکلئوتید مکمل بین پرایمرها) که با اتصال پرایمر به الگوی DNA تداخل میکند. باید همسانی درون پرایمر یا بین پرایمرها را تا حد ممکن پایین نگه داشته شود. تا به امروز برنامههای زیادی برای طراحی پرایمر در ژنتیک ایجاد شده است. در اینجا برخی از برنامههای تحت وب و مستقل را معرفی میکنیم که برای استفاده عمومی رایگان هستند.
- Primer3. این وب سایت که متعلق به دانشکده پزشکی ماساچوست آمریکا است دارای یک برنامه طراحی آغازگر PCR بسیار قدرتمند است که به شما امکان کنترل قابل توجهی بر ماهیت پرایمرها را میدهد، از جمله اندازه محصول مورد نظر، اندازه پرایمر در ژنتیک و دامنه TM و وجود یا عدم وجود گیره ’3-GC.
- GeneFisher. که یک ابزار طراحی آغازگر PCR تعاملی متعلق به دانشگاه بیلفلد آلمان است و یک سایت بسیار خوب که امکان کنترل عالی بر طراحی پرایمر در ژنتیک را دارد.
- Primer3Plus. یک رابط وب جدید بهبود یافته برای برنامه محبوب پرایمر در ژنتیک Primer3 است.
- BiSearch. یک ابزار طراحی و جستجوی پرایمر در ژنتیک که ابزار مفیدی جهت طراحی آغازگر برای هر الگوی DNA و به ویژه برای ژنومهای تحت درمان با بیسولفیت است. ابزار ePCR امکان شناسایی سریع جایگاههای اشتباه و محصولات جایگزین PCR را در کتابخانههای cDNA و ژنومهای تحت درمان با بیسولفیت فراهم میکند.

- Primer-BLAST. در وب سایت NCBI برای کمک به کاربران در ساخت آغازگرهایی که خاص الگوی ورودی PCR هستند، توسعه داده شد. از Primer3 برای طراحی آغازگرهای PCR استفاده میکند و سپس آنها را برای جستجوی BLAST در پایگاه داده انتخاب شده توسط کاربر ارسال میکند. سپس نتایج بلاست به طور خودکار تجزیه و تحلیل میشود تا از جفتهای پرایمر در ژنتیک که میتوانند باعث تکثیر اهداف دیگری غیر از الگوی ورودی شوند، جلوگیری شود.
- MFEprimer. به کاربران اجازه میدهد تا به سرعت و به راحتی اختصاصیت آغازگر را در برابر DNA ژنومی و RNA پیام رسان در پایگاه دادههای توالی DNA مکمل بررسی کنند. این سرور از یک الگوریتم شاخص k-mer برای تسریع روند جستجو در سایتهای اتصال پرایمر در ژنتیک استفاده کرده و از ترمودینامیک برای ارزیابی ثبات اتصال بین هر آغازگر و الگوی DNA آن استفاده میکند.
- PrimerDesign-M. شامل چندین گزینه برای طراحی پرایمرهای متعدد است و به محققان اجازه میدهد تا بطور موثر آغازگرهای حرکت کننده در طول رشته هدف را که اهداف طولانی DNA را پوشش میدهند، طراحی کنند مانند کل ژنوم های HIV-1، و بهینهسازی آغازگرها به طور همزمان با تنوع ژنتیکی در چندین ترازبندی. این ابزار محدودیتهای طراحی تجربی انجام شده توسط کاربر را ندارند. PrimerDesign-M همچنین میتواند آغازگرهایی را طراحی کند که شامل بارکد DNA باشند و پرایمر - دایمر را به حداقل برسانند. PrimerDesign-M آغازگرهای بهینه را برای اهداف DNA بسیار متغیر پیدا میکند و با پیشنهاد طرحهای جایگزین برای انطباق با شرایط آزمایشی، انعطاف پذیری طراحی پرایمر در ژنتیک را تسهیل میکند.
- RF-cloning. یا «شبیه سازی بدون محدودیت» (Restriction-free cloning) یک فناوری مبتنی بر PCR است که در فرآیند «جهش زایی تغییر سریع» (QuikChange™ mutagenesis) گسترش مییابد که در ابتدا توسط Stratagene در اواسط دهه 1990 رواج یافته و اجازه میدهد اساساً هر توالی را به هر پلاسمید در هر جایگاهی وارد کنید.

- primers4clades. خط لولهای برای طراحی آغازگرهای PCR جهت تکثیر توالیهای جدید از DNA متاژنومی یا ارگانیسمهای ناشناخته متعلق به تبارهای فیلوژنتیک مشخص شده توسط کاربر است. این ابزار یک استراتژی گسترده CODEHOP را بر اساس چندین ترازبندی DNA و پروتئین ژنهای کدکننده و خصوصیات ترمودینامیکی جفتهای الیگونوکلئوتیدی و همچنین محتوای اطلاعات فیلوژنتیک در آمپلیکونهای پیش بینی شده، محاسبه شده با مقادیر برگرفته از حداکثر شباهت را اجرا میکند. درختهای فیلوژنیکی که بر روی صفحه نمایش داده میشوند، هدف قرار دادن پرایمرها به تیغههای تعاملی را آسان میکنند.
- TaxMan. آمپلیکونهای rRNA و نوع گونهها در این ابزار قابل بررسی هستند. در تجزیه و تحلیل میکروبیوم، اغلب از پایگاه دادههای ژن rRNA برای اختصاص اسامی تاکسونومیک به توالیهای خواندنی استفاده میشود. سرور TaxMan تجزیه و تحلیل توزیع تاکسونومیک توالیهای خوانده شده را از دو طریق تسهیل میکند. ابتدا میتوانید بررسی کنید که چه اسامی تاکسونومیکی به توالیهای تولید شده توسط پرایمرهای شما اختصاص داده میشوند و چه گونههایی از دست خواهید داد. دوم، توالیهای آمپلیکون تولید شده با اجداد را در FASTA میتوان مطابقت داد. این میتواند منجر به یک تجزیه و تحلیل بسیار کارآمدتر با توجه به زمان اجرا و استفاده از حافظه مورد استفاده شود، زیرا توالیهای آمپلیکون بطور قابل توجهی کوتاهتر از توالیهای ژن rRNA با طول کامل هستند. به علاوه از طریق این ابزار میتوانید یک فایل دودمان بارگیری کنید که شامل شمارش تمام گونهها برای پرایمر در ژنتیک و مرجع استفاده شده است.

ابزار بررسی ساختاری پرایمر در ژنتیک
علاوه بر ابزارهای بیان شده در طراحی پرایمرهای مختلف ابزارهای مفید دیگر و وب سایتهای فعالی در زمینه بررسی ساختارهای شیمیایی پرایمر در ژنتیک و الیگونوکلئوتیدها و همچنین پیش بینی ایجاد ساختارهای ثانویه در روند پژوهش نیز در دسترس است که در ادامه به بررسی آنها میپردازیم.

- NetPrimer. این سایت با Tm، خواص ترمودینامیکی و پیشبینی ساختارهای سنجاق سری و پرایمر - دایمر را فراهم میکند، اما بارگیری برنامه مدتی طول میکشد.
- dnaMATE. در این ابزار محاسبه Tm برای توالی DNA کوتاه (16 تا 30 نوکلئوتید) با استفاده از یک روش ادغام شده بر اساس سه جدول مختلف ترمودینامیکی صورت میگیرد. مقدار Tm برآوردی قوی و دقیق از دمای ذوب برای توالیهای کوتاه کاربردی DNA در زیست شناسی مولکولی است.
- OligoCalc. یک محاسبه کننده آنلاین خواص الیگونوکلئوتید است.
- OligoAnalyzer 3.1.
- Mongo Oligo Mass Calculator v2.06.
- OligoEvaluator.
- Oligo Calculation Tool. یک ابزار محاسبه کننده الیگونوکلئوتیدها است که به کاربر اجازه اصلاح توالی را نیز میدهد.
ابزار طراحی پرایمر بر اساس توالی پروتئین
اگر شما توالی پروتئینی در دست دارید و توالی DNA آن را میخواهید یکی از بهترین وب سایتها «ترجمه معکوس پروتئین به DNA» یا وب سایت (Protein to DNA reverse translation or Reverse Translation) که از ابزارهای دستکاری توالی هستند. اگر شما علاقه مند به تغییر یک اسید آمینه خاص به آمینواسید دیگری هستید، باید از Primaclade استفاده کنید.

ابزار طراحی پرایمر PCR و کلونینگ
AMUSER یا (Automated DNA Modifications with USER cloning) ابزاری است که طراحی سریع و آسان پرایمرهای PCR را برای مهندسی DNA مبتنی بر USER cloning بهینهسازی کرده است. USER cloning یک روش سریع و همهکاره برای مهندسی DNA پلاسمید است. این ابزار وب سرور طراحی آغازگرهای بهینه PCR را برای چندین برنامه مبتنی بر USER cloning به طور خودکار انجام میدهد. با طراحی آغازگرهای بهینه PCR برای تغییرات ژنتیکی مطلوب، مونتاژ DNA و مقدمه تقریباً هر نوع جهشزایی هدایت شده مکانی را به کار میگیرد.
The PCR Suite یک وب سایت دیگر بوده و این مجموعه از چهار برنامه مبتنی بر Primer3 برای طراحی آغازگر ژنومی است. که همه آنها کنترل قابل توجهی بر خصوصیات آغازگر دارند و شامل مجموعههای زیر هستند.
- Overlapping_Primers. چندین محصول PCR همپوشان را در یک توالی ایجاد میکند.
- Genomic_Primers. آغازگرهای اطراف اگزونها را در توالی ژنومی طراحی میکند. تمام آنچه نیاز دارید یک فایل GenBank حاوی ژن مورد نظر شماست.
- SNP_Primers. آغازگرها را در اطراف هر SNP در یک فایل GenBank طراحی میکند.
- cDNA_Primers. آغازگرها را در اطراف چارچوبهای خواندنی بازها طراحی میکند. که در اینجا نیز به سادگی میتوانید یک فایل GenBank حاوی ژنهای مورد نظر خود را بارگذاری کنید.
هنگامی که آماده راه اندازی واکنش PCR خود هستید، به PCR Box Titration Calculator مراجعه کنید، برای فهمیدن مقدار هر معرف برای استفاده در یک تیتراسیون جعبه دو بعدی برای PCR و برای واکنشهای PCR استاندارد، میزان حجم را تنظیم کرده و شماره سطر و ستون را به 1 تغییر دهید، روی موارد بالا یا پایین و تمام کلیک کنید. همچنین از PCR Reaction Mixture Setup نیز میتوانید استفاده کنید که یک وب سایت بسیار خوب برای این مرحله است. به علاوه از ابزار PCR Optimization نیز میتوان استفاده کرد که دارای امکانات بسیار خوبی در این زمینه است.
برای ارائه پرایمر در توالی DNA میتوان از Sequence Extractor استفاده کرد که یک نقشه محدود قابل کلیک و یک آغازگر PCR از یک توالی DNA ایجاد میکند (قالبهای پذیرفته شده عبارتند از: raw ، GenBank ، EMBL و FASTA) که کنترل زیادی بر خروجی ارائه میدهد. همچنین در آن ترجمه پروتئین و مرزهای اینترون / اگزون نیز نشان داده شده است. برای ساخت سازههای DNA به صورت کامپیوتری (in-silico) از Sequence Extractor استفاده کنید.

ابزار طراحی پرایمر همپوشان
دو وب سایت ارائه دهنده نرم افزار بر اساس برنامه Primer3 برای طراحی مجموعههای جفت آغازگر PCR با هم تداخل دارند که شامل Multiple Primer Design with Primer و Overlapping Primersets هستند. علاوه بر این دو مورد وب سایت PHUSER یا Primer Help for USER «همجوشی واکنش خاص اوراسیل» (USER) یک تکنیک اخیراً توسعه یافته است که امکان جمع آوری قطعات DNA متعدد را در چند مرحله ساده فراهم میکند. PHUSER با طراحی سریع و آسان آغازگرهای بهینهسازی شده PCR، از ادغام صحیح قطعات در یک پلاسمید حاوی یک کاست قابل تنظیم برای کاربر اطمینان حاصل میکند.
PHUSER همچنین از برآمدگیهای یکسان جلوگیری کرده و بدین روش ترتیب صحیح مونتاژ قطعات DNA را تضمین میکند. تمام آغازگرهای ممکن از نظر محتوای GC، وجود گیره GC در انتهای ’3، خطر تشکیل پرایمر - دایمر، خطر ساختارهای ثانویه درون پرایمر و وجود کشیدگیهای polyN به صورت جداگانه مورد تجزیه و تحلیل قرار میگیرند. ابزار دیگر Primerize بوده که یک سرور وب برای طراحی و مونتاژ پرایمرهای توالی DNA در PCR است. Primerize برای کاهش اتصالات غیر اختصاصی اولیه آغازگرها بهینه سازی شده است، برای توالیهای ثابت مشکلات RNA طراحی شده است و آزمایشهای گسترده و دقیق را پشت سر گذاشته است.
ابزار طراحی siRNA ها
«RNA تداخل کننده کوچک» (siRNA) تخریب توالی خاص mRNA همولوگ را هدایت میکند، بنابراین سلولهای مجزا ایجاد میکند. ابزار طراحی siRNA یک ژن هدف را برای توالی کاندیداهای siRNA که قوانین قابل تنظیم توسط کاربر را برآورده کرده، اسکن میکند. انواع مختلفی از سرورها برای این عملکرد وجود دارند که شامل موارد زیر هستند.
- siRNA Target Finder.

- siRNA Design Software. ابزارهای طراحی موجود از جمله موارد ذکر شده در بالا را مقایسه میکند. آنها همچنین سعی دارند اصول MPI و ابزارهای موجود را با استفاده از الگوریتمی که میتواند siRNA های بی اثر را فیلتر کند، بهبود بخشد، این الگوریتم بر اساس برخی مشاهدات جدید در مورد ساختار ثانویه است.
- OligoWalk. یک سرور آنلاین است که ویژگیهای ترمودینامیکی هیبریدسازی sense - antisense را محاسبه میکند. این ابزار تغییرات انرژی آزاد اتصال الیگونوکلئوتیدها به RNA هدف را پیشبینی میکند. میتوان از آن برای طراحی siRNA کارآمد استفاده کرد که یک توالی mRNA داده شده را هدف قرار میدهد.
- VIRsiRNApred. یک سرور پیش بینی اثربخشی siRNA ویروسی انسان است.
- Dicer-substrate siRNAs. (DsiRNA ها) RNA های دابلکس 27 زیرواحدی سنتزی شده شیمیایی هستند که قدرت تداخل RNA را در مقایسه با siRNA های سنتی افزایش میدهند.
- pssRNAit. ابزار طراحی RNAi siRNA های گیاهی موثر و خاص با ارزیابی ژن خارج هدف از نظر ژنوم هستند.
- DSIR. ابزاری برای طراحی هدف siRNA (19 یا 21 نوکليوتیدی) و shRNA است.
- Imgenex siRNA. برنامه بازیابی ساخته شده برای انتخاب الیگونوکلئوتیدهای DNA رمزگذار siRNA است که میتواند در یکی از وکتورهای pSuppressor کلون شود. توالی ورودی را میتوان از طریق الحاق Genbank یا توالی تهیه شده توسط محقق به طور مستقیم در دسترس قرار داد.
- siDRM. پیاده سازی مجموعه قانون DRM برای انتخاب siRNA های موثر است. پژوهشگران با استفاده از رویکرد ادغام قاعده انقطاع (DRM) در یک مجموعه داده بزرگ و متنوع که از siRecords گردآوری شده است، تجزیه و تحلیل به روز شدهای را انجام داده و مجموعه قوانین حاصل را در siDRM، یک ابزار جدید طراحی siRNA آنلاین، پیاده سازی کردهاند. siDRM همچنین چند مجموعه قانون با حساسیت بالا و مجموعه قوانین سریع، پیوند یافته به siRecords را اجرا میکند و از چندین فیلتر برای بررسی اثرات مخرب ناخواسته، از جمله پاسخهای ایمنی ذاتی، اثرات سمی سلول و فعالیتهای خارج از هدف در انتخاب siRNA ها استفاده میکند.
- siMAX siRNA Design Tool. یک نرم افزار اختصاصی است که برای کمک در انتخاب مناسب ترین siRNA ژن مورد علاقه شما طراحی شده است.
- shRNA Designer. از این برنامه برای طراحی الیگوهای shRNA که با وکتورهای SORT-A / B / C سازگار هستند استفاده میشود. این ابزار طراحی، اهدافی را با بیشترین شانس برای از بین بردن ژن شما فراهم میکند. لطفا توجه داشته باشید، فقط یک الیگو به عنوان پالیندرومیک طراحی شده است.
- siDESIGN Center. یک ابزار طراحی پیشرفته و کاربر پسند siRNA است، که به طور قابل توجهی احتمال شناسایی siRNA عملکردی را بهبود میبخشد. گزینههای هم نوع برای افزایش ویژگی هدف و انطباق طرحهای siRNA برای طراحی آزمایشگاهی پیچیدهتر در دسترس هستند.

ابزار طراحی پرایمر Real-time PCR
همانطور که از نام آنها پیداست این ابزارهای تحت وب برای طراحی پرایمرهای تکنیک Real-time PCR استفاده میشوند و با سایر ابزارهای بیان شده تفاوتهای اندکی دارند. در ادامه برخی از بهترین ابزارهای طراحی پرایمر در ژنتیک برای تکنیک Real-time PCR را توضیح میدهیم.
- RealTimeDesign. این ابزار رایگان بوده اما نیاز به ثبت نام دارد.
- GenScript Real-time PCR (TaqMan) Primer Design. میتواند دامنه اندازه امپلیکون PCR را و همچنین TM (دمای ذوب) برای پرایمرها و پروبها و ارگانیسم سفارشی کرد. همچنین میتوانید تصمیم بگیرید که چند مجموعه پرایمر/پروب میخواهید این ابزار به شما بازگرداند. استفاده از شماره الحاقی GenBank به عنوان الگو امکانپذیر است.
- QuantPrime. یک برنامه انعطافپذیر برای طراحی آغازگر قابل اعتماد برای استفاده در آزمایشهای qPCR بزرگتر است. چارچوب انعطافپذیر همچنین برای استفاده ساده در سایر کاربردهای کمی، مانند طراحی پروب هیدرولیز برای qPCR و طراحی پروب الیگونوکلئوتید برای هیبریدسازی کمی in situ، باز است.
- PrimerQuest.
ابزار طراحی پرایمر جهش زا
WatCut یک الیگونوکلئوتید میگیرد و جهشهای خاموش را در مکانهای محدود کننده احتمالی ایجاد میکند، به طوری که توالی اسید آمینه پروتئین تغییر نکرده است. ابزار دیگر PrimerX است که میتواند مورد استفاده قرار گیرد تا به طور خودکار طراحی آغازگرهای جهشزا برای جهش زایی هدایت شده مکانی را انجام دهد این ابزار دارای دو حالت است: الف) طراحی آغازگر بر اساس توالی DNA و ب) طراحی پرایمر مبتنی بر توالی پروتئین. ابزار مورد استفاده بعدی Primerize-2D است که برای سرعت بخشیدن به سنتز کتابخانههای بزرگ جهشهای مورد نظر از طریق طراحی و سازماندهی کارآمد آغازگرها طراحی شده است.









