



یوکاریوت چیست؟ – به زبان ساده + تعریف و مثال
موجودات یوکاریوتی بسیار پیچیدهتر از انواع پروکاریوتی هستند. سلولهای آنها بزرگتر بوده و دارای اندامکهای متصل به غشاء مانند هسته، میتوکندری و کلروپلاست هستند. سلولهای بدن انسان یوکاریوتی است. مشخصه اصلی یوکاریوتها حضور هسته در داخل سلول است. در این مطلب به بررسی یوکاریوتها، تعریف سلول یوکاریوتی و ساختار آن، انواع یوکاریوتها و طبقهبندی آنها میپردازیم.
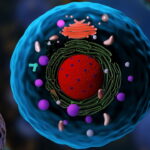
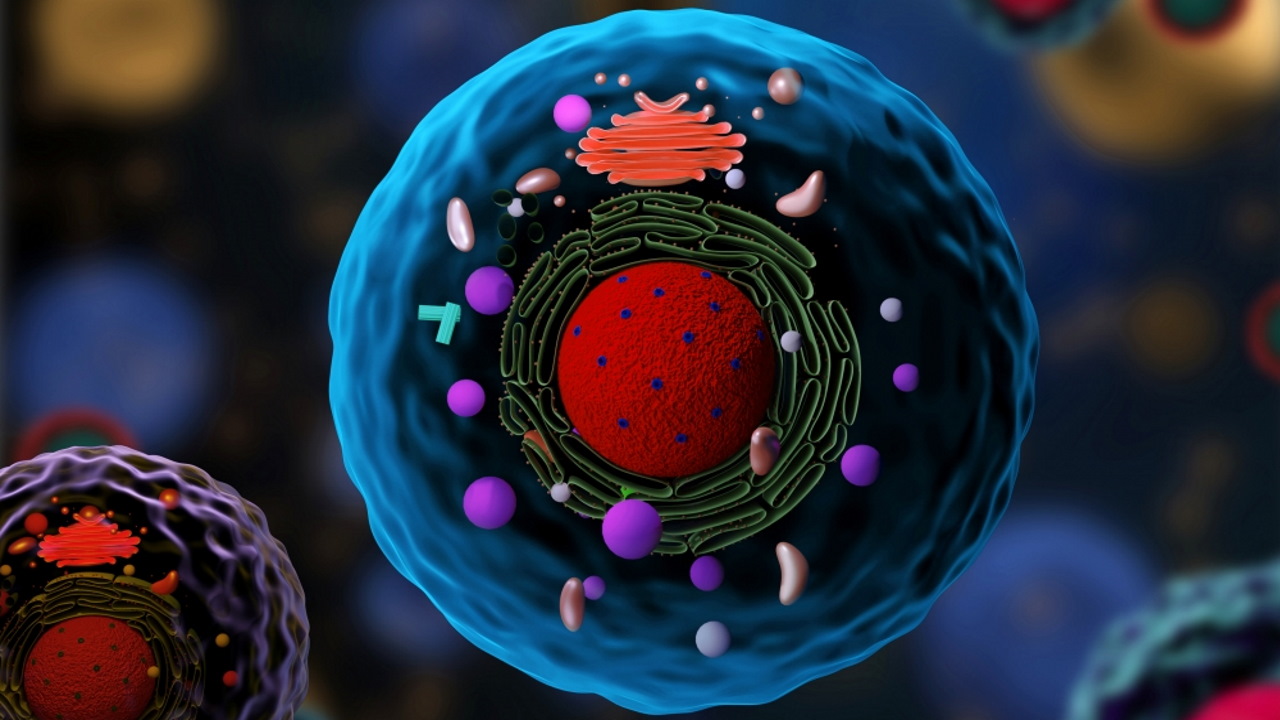


یوکاریوت چیست؟
یوکاریوت به هر کدام از موجودات تکسلولی یا چندسلولی اطلاق میشود که سلول آنها دارای یک هسته مجزا و متصل به غشاء است. موجوداتی مانند حیوانات، گیاهان، قارچها و آغازیان نمونههایی از یوکاریوتها هستند زیرا سلولهای آنها در ساختارهای تقسیمبندیشده به نام اندامکها و هسته سازماندهی شدهاند. وجود هسته مجزا که درون غشاها قرار گرفته است، یوکاریوتها را از پروکاریوتها متمایز میکند. یوکاریوتها همچنین به داشتن اندامکهای سیتوپلاسمی جدا از هسته مانند میتوکندری، کلروپلاست و اجسام گلژی معروف هستند.
یوکاریوت به هر ارگانیسمی گفته میشود که عمدتاً توسط سلولی با یک یا چند هسته در طول زندگی خود مشخص میشود، برخلاف پروکاریوتی که دارای سلولی فاقد هسته مشخص و فقط دارای ناحیه نوکلوئیدی است.

تکامل یوکاریوت ها
سلولهای یوکاریوتی از سلولهای پروکاریوتی بین 1/6 تا 2/7 میلیارد سال پیش تکامل یافتهاند. امروزه همه موجودات پیچیده و بیشتر موجودات چندسلولی یوکاریوت هستند و این تکامل را به رویدادی بزرگ در تاریخ حیات روی زمین تبدیل کرده است. حدود 75 دودمان جداگانه از یوکاریوتها وجود دارد که بیشتر آنها به پروتیستها (آغازیان) تبدیل شدهاند. یوکاریوتها بیشتر با ارگانیسمهای تکسلولی که گاهی در شرایط شدید مانند چشمههای آب گرم یافت میشوند، بیشتر از باکتریها مرتبط هستند.
سلول های یوکاریوتی اندامکهای خاصی را ایجاد کردند که ساختارهایی در درون سلول هستند که وظیفه خاصی را انجام میدهند. این اندامک ها عبارتند از میتوکندریها که انرژی میسازند، کلروپلاستها که در گیاهان یافت میشوند و از نور و دیاکسیدکربن غذا میسازند و شبکه آندوپلاسمی که پروتئینها را دستهبندی و بستهبندی میکند. برخی از اندامکها، مانند میتوکندری و کلروپلاست، ممکن است زمانی تکامل یافته باشند که باکتریهای آزاد به درون سلولها جذب شدهاند. بر اساس این نظریه، باکتریها و سلولها رابطه همزیستی داشتند که در آن هر یک از وجود دیگری سود میبرد.
نظریه درون همزیستی
با گذشت زمان، این باکتریها اندامکهایی را تشکیل دادند که درون سلولهای یوکاریوتی که امروزه دیده میشوند، ادغام شده و به بخشی ضروری از سلول یوکاریوتی تبدیل شدند. میتوکندریها دارای DNA هستند که جدا از DNA کروموزومی موجود در هسته سلول است. با این حال، نظریه دیگر این است که مقادیر کمی از DNA موجود در سلول در غشای سلولی پوشانده شده و به اندامکهایی مانند میتوکندری تبدیل شده است.
نظریههای مربوط به میتوکندریها در سلولی که قبلاً تقریباً یوکاریوتی بوده وجود دارد به این ترتیب که این سلولها با یک هسته ظاهر شده و به عنوان مدلهای خودزا شناخته میشوند. با این حال، نظریه دیگری پیشنهاد میکند که سلولهای یوکاریوتی زمانی تکامل یافتند که یک آرکئون و یک باکتری با هم ادغام شدند و سلول جدیدی را تشکیل دادند. این به عنوان مدل کایمریک شناخته میشود. برخی از یوکاریوتها به صورت غیرجنسی تولیدمثل میکنند، در حالی که برخی دیگر از طریق جنسی تولید مثل میکنند. توسعه تولیدمثل جنسی یکی دیگر از ویژگیهای تعیینکننده در تکامل یوکاریوتها است.

اعتقاد بر این است که اجداد مشترک همه یوکاریوتها از طریق جنسی تولید مثل میکردند و یوکاریوتهای غیرجنسی (مانند برخی از آمیبها) این روش تولید مثل را از اجدادی که جنسی بود تکامل دادند. پروکاریوتها فقط به صورت غیرجنسی تولید مثل می کنند. ژنها را میتوان از طریق انتقال افقی ژن بین افراد مبادله کرد، اما این تولید مثل جنسی نیست.
پروکاریوت و یوکاریوت
پروکاریوتها ارگانیسمهایی هستند که با فقدان هسته و سایر ساختارهای سیتوپلاسمی متصل به غشاء مشخص میشوند. آنها به طور قابل توجهی کوچکتر از یوکاریوتها هستند و همچنین نسبت سطح به حجم بیشتری داشته، بنابراین نرخ متابولیک بالاتری دارند. نمونههایی از پروکاریوتها، یوباکتریها و آرکئاها هستند.
یوکاریوتها دارای هستهای هستند که حاوی DNA هستهای است. هسته دارای غشای دولایه لیپیدی با منافذ هستهای است. DNAهای داخل هسته با پروتئینهای هیستون کمپلکس شده و کروماتین را تشکیل میدهند. کروموزومها رشتههای خطی DNA بوده اما در مقابل کروموزومهای پروکاریوتها عمدتاً دایرهای هستند. هم یوکاریوتها و هم پروکاریوتها اطلاعات ژنتیکی ذخیره شده در ژنها دارند. منبع اصلی انرژی متابولیک آنها ATP است. هر دوی آنها همچنین دارای ریبوزومهایی هستند که در سنتز پروتئین کمک میکنند.

با این حال، ریبوزومهای یوکاریوتها 80S بوده اما در پروکاریوتها، ریبوزومها 70S هستند. ریبوزومهای پروکاریوتی و یوکاریوتی هر دو از دو زیرواحد ریبوزومی ساخته شدهاند. ریبوزوم پروکاریوتی (70S) از زیرواحدهای 50S (زیرواحد بزرگ) و 30S (زیرواحد کوچک) تشکیل شده است. ریبوزوم یوکاریوتی (80S) از 60S (زیرواحد بزرگ) و 40S (زیرواحد کوچک) تشکیل شده است. توجه داشته باشید واحدهای S با هم جمع نمیشوند، زیرا آنها معیارهای نرخ رسوب را نشان میدهند و نشاندهنده جرم نیستند.
سلول یوکاریوتی چیست؟
سلول یوکاریوتی دارای چندین ساختار متصل به غشاء است که در سیتوپلاسم پراکنده شده و اندامکها نامیده میشوند. اندامکهایی که معمولا در داخل سلول یوکاریوتی یافت میشوند عبارتند از: هسته، شبکه آندوپلاسمی، دستگاه گلژی، میتوکندری و پلاستید. سایر ساختارهای سیتوپلاسمی عبارتند از اسکلت سلولی، اجسام وزیکولی و بیومولکولها. سلولهای یوکاریوتی معمولاً بسیار بزرگتر از سلولهای پروکاریوتها هستند و حجمی حدود 10000 برابر بیشتر از سلولهای پروکاریوتی دارند. DNA یوکاریوتی به چند دسته خطی به نام کروموزوم تقسیم شده که در طی تقسیم هستهای توسط دوک میکرولولهای از هم جدا میشوند.
سلول یوکاریوت گیاهی
سلولهای گیاهی یوکاریوتی هستند که در چندین عامل اساسی با سایر موجودات یوکاریوتی تفاوت دارند. سلولهای گیاهی و حیوانی هر دو حاوی هسته همراه با اندامکهای مشابه هستند. یکی از جنبههای متمایز سلول گیاهی وجود دیواره سلولی در خارج از غشای سلولی و همچنین اندامک کلروپلاست است. سلول گیاهی مستطیل شکل و نسبتاً بزرگتر از سلول حیوانی است. اگرچه سلولهای گیاهی و حیوانی یوکاریوتی هستند و اندامکهای سلولی مشترکی دارند، سلولهای گیاهی در مقایسه با سلولهای حیوانی کاملاً متمایز هستند زیرا عملکردهای متفاوتی دارند.

ساختار سلول گیاهی
درست مانند اندامهای مختلف بدن، ساختار سلولهای گیاهی شامل اجزای مختلفی است که به نام اندامکهای سلولی شناخته میشوند که عملکردهای مختلفی را برای حفظ خود انجام میدهند. این اندامکها عبارتند از:
- دیواره سلولی: این لایه سفت و سخت است که از پلی ساکاریدهای سلولز، پکتین و همیسلولز تشکیل شده است. این بخش خارج از غشای سلولی قرار دارد. همچنین شامل گلیکوپروتئینها و پلیمرهایی مانند لیگنین، کوتین یا سوبرین است. وظیفه اصلی دیواره سلولی محافظت و حمایت ساختاری از سلول است. تشکیل دیواره سلولی توسط میکروتوبولها هدایت میشود و در سلول گیاهی از سه لایه به نامهای اولیه، ثانویه و تیغه میانی تشکیل شده است.
- غشای سلول: غشا به صورت ساختاری دولایه فسفولیپیدی است که در جدا کردن سلول از محیط اطرافش نقش اصلی را دارد. غشای سلولی همچنین نقش مهمی در تنظیم ورود و خروج مواد خاص در داخل سلول دارد.
- هسته: هسته، ساختاری متصل به غشاء است که فقط در سلولهای یوکاریوتی وجود دارد. عملکرد حیاتی هسته ذخیره DNA یا اطلاعات ارثی مورد نیاز برای تقسیم سلولی، متابولیسم و رشد است.
- کلروپلاست: این اندامک طویل بوده و توسط غشای فسفولیپیدی محصور شده است. کلروپلاست شبیه یک دیسک است و استروما مایع سیال درون کلروپلاست است که حاوی DNA دایرهای است. هر کلروپلاست حاوی رنگدانه سبز به نام کلروفیل است که برای فرآیند فتوسنتز لازم است. کلروفیل انرژی نور خورشید را جذب میکند و از آن برای تبدیل دیاکسیدکربن و آب به گلوکز استفاده میکند.
- لوکوپلاستها: آنها در بافت غیر فتوسنتزی گیاهان یافت میشوند و برای ذخیره پروتئین، لیپید و نشاسته استفاده میشوند.
- واکوئل مرکزی: حدود 30 درصد از حجم سلول را در سلول گیاهی بالغ اشغال میکند. تونوپلاست غشایی است که واکوئل مرکزی را احاطه کرده است. عملکرد حیاتی واکوئل مرکزی جدا از ذخیرهسازی، حفظ فشار تورژسانس در برابر دیواره سلولی است. محتویان داخل واکوئل مرکزی از شیره سلولی تشکیل شده که ترکیبی از نمکها، آنزیمها و سایر مواد است.
- دستگاه گلژی: آنها در تمام سلولهای یوکاریوتی یافت میشوند که در توزیع ماکرومولکولهای سنتز شده در قسمتهای مختلف سلول نقش دارند.
- ریبوزوم: این ساختارها کوچکترین اندامکهای متصل به غشاء هستند که شامل RNA و پروتئین هستند. آنها مکانهایی برای سنتز پروتئین بوده، از این رو به عنوان کارخانههای پروتئینسازی سلول نیز شناخته میشوند.
- میتوکندری: آنها اندامکهای دو غشایی هستند که در سیتوپلاسم تمام سلولهای یوکاریوتی یافت میشوند. آنها انرژی را با شکستن مولکولهای کربوهیدرات و قند تامین میکنند، از این رو به آنها «نیروگاه سلول» نیز میگویند.
- لیزوزوم: لیزوزومها کیسههای خودکشی نامیده میشوند زیرا آنزیمهای گوارشی را در یک غشای محصور نگه میدارند. آنها عملکرد دفع زبالههای سلولی را با هضم اندامکهای فرسوده، ذرات غذا و اجسام خارجی در سلول انجام میدهند. در گیاهان، نقش لیزوزومها توسط واکوئلها انجام میشود.

انواع سلول های گیاهی
سلولهای گیاه بالغ و عالی برای انجام برخی عملکردهای حیاتی که برای بقای آنها ضروری است، بسته به عملکرد خاص خود تخصصی میشوند. تعداد کمی از سلولهای گیاهی در حمل و نقل مواد مغذی و آب نقش داشته، در حالی که سلولهای دیگر برای ذخیره غذا نقش دارند. سلولهای گیاهی تخصصی شامل سلولهای پارانشیم، سلولهای اسکلرانشیم، سلولهای کلانشیم، سلولهای آوند چوبی و سلولهای آبکش هستند. در زیر به برخی از انواع مختلف سلولهای گیاهی اشاره کردهایم:
- سلولهای کلانشیمی: آنها سلولهای سفت و سختی هستند که نقش اصلی را در حمایت از گیاهان دارند.
- سلول اسکلرانشیم: این سلولها در مقایسه با سلولهای کلانشیمی سفتتر هستند و این به دلیل وجود عامل سختکننده است. این سلولها معمولاً در همه ریشههای گیاهان یافت میشوند و عمدتاً در حمایت از گیاهان نقش دارند.
- سلول پارانشیم: سلولهای پارانشیم در همه گیاهان نقش بسزایی دارند. آنها سلولهای زنده گیاهان هستند که در تولید برگها نقش دارند.
- سلول زایلم: آنها همچنین در تبادل گازها، تولید غذا، ذخیره محصولات ارگانیک و متابولیسم سلولی نقش دارند. این سلولها معمولاً انعطافپذیرتر از سایرین بوده، زیرا نازکتر هستند.
- سلول آبکش: سلولهای آبکش دیگر سلولهای انتقالدهنده در گیاهان آوندی هستند. آنها غذای تهیه شده توسط برگها را به قسمتهای مختلف گیاهان منتقل میکنند.
عملکرد سلول گیاهی
سلولهای گیاهی اجزای سازنده گیاهان هستند. فتوسنتز عملکرد اصلی سلولهای گیاهی است. فتوسنتز در کلروپلاستهای سلول گیاهی اتفاق میافتد. این فرآیند تهیه غذا توسط گیاهان با استفاده از نور خورشید، دیاکسیدکربن و آب است. در این فرآیند انرژی به شکل ATP تولید میشود. چند سلول گیاهی به انتقال آب و مواد مغذی از ریشه و برگ به قسمتهای مختلف گیاهان کمک میکنند.

سلول یوکاریوت جانوری
سلولهای حیوانی حاوی اندامکهای متنوعی هستند که به آنها کمک میکند تا وظایف خود را انجام دهند. سلولهای حیوانی همانند آنچه در باکتریها نیز وجود دارد دارای غشای پلاسمایی، سیتوپلاسم و DNA هستند. با این حال، متوجه خواهید شد که داخل و خارج سلولهای حیوانی کاملاً متفاوت از باکتریها به نظر میرسد. برای مثال، سلولهای حیوانی دیواره سلولی ندارند. در عوض، آنها اسکلت سلولی دارند که شبکهای از رشتههای متشکل از پروتئین است. اسکلت سلولی پشتیبانی و حمل و نقل داخلی را برای سلول فراهم میکند. علاوه بر این، بیشتر سلولهای جانوری دارای هسته هستند، اندامک خاصی که DNA را ذخیره میکند. DNA درون هسته معمولاً به صورت رشتههایی به نام کروموزوم سازماندهی میشود.
ساختار سلول یوکاریوت جانوری
سیتوپلاسم سلولهای حیوانی پر از اندامکهای مختلفی است که به زنده ماندن و تولید مثل سلولها کمک میکند. در اینجا برخی از اندامکهای اصلی ساکن در سیتوپلاسم و عملکرد آنها آورده شده است:
- سانتروزوم: سانتریولها و مواد پیشسانتریولی داخل سلول در تقسیم سلولی و ساختن میکروتوبولها نقش دارند.
- دستگاه گلژی: پروتئینها را اصلاح و مرتب کرده و در وزیکولهای انتقالی بستهبندی میکند، بنابراین آنها میتوانند به جای دیگری در سلول بروند یا ترشح شوند.
- لیزوزوم و پراکسیزوم: به حذف مواد زائد، شکستن ترکیبات سمی و بازیافت ساختارهای سلولی کمک میکند.
- میتوکندری: انرژی تولید میکند.
- ریبوزوم: سنتز پروتئین را انجام میدهد.
- شبکه اندوپلاسمی زبر: پیوسته با لایه بیرونی پوشش هستهای و دارای ریبوزومهای تعبیه شده بر روی غشای خارجی است. به انتقال مواد در داخل سلول کمک میکند. پروتئینهای تازه ساخته شده را برای انتقال توسط وزیکولها جدا میکند.
- شبکه اندوپلاسمی صاف: جدا از غشای هسته، اما پیوسته با شبکه آندوپلاسمی زبر بوده ولی حاوی ریبوزوم نیست. این اندامک محل سنتز لیپید، متابولیسم کربوهیدرات و سمزدایی بوده و به انتقال مواد در داخل سلول کمک میکند.
- وزیکول: کیسههای غشایی کوچک که مواد را در داخل سلول حمل میکنند. میتواند با غشای سلول ترکیب شود تا محتویات را آزاد کند.

سلول یوکاریوت قارچی
قارچها تجزیهکنندههای هتروتروف با دیواره سلولی ضخیم تکسلولی یا چندسلولی هستند که مواد پوسیده را میخورند و رشتههای در هم تنیدهای را ایجاد میکنند. دیواره سلولی قارچی سفت و سخت بوده و حاوی پلیساکاریدهای پیچیدهای به نام کیتین (برای افزایش استحکام ساختاری) و گلوکان است. قارچها میتوانند تکسلولی، چندسلولی یا دوشکلی باشند که بسته به شرایط محیطی، تکسلولی یا چندسلولی هستند. قارچها در مرحله رویشی مورفولوژیکی از مجموعهای از هیفهای باریک و نخمانند تشکیل شدهاند، در حالی که مرحله تولید مثل معمولاً آشکارتر است.
قارچها دوست دارند در محیط مرطوب و کمی اسیدی باشند. آنها میتوانند با یا بدون نور یا اکسیژن رشد کنند. قارچها هتروتروفهای ساپروفیت هستند زیرا از مواد آلی مرده یا در حال تجزیه به عنوان منبع کربن استفاده میکنند.
ساختار سلول قارچی
قارچها یوکاریوت هستند و دارای سازمان سلولی پیچیدهای هستند. سلولهای قارچی به عنوان یوکاریوتها حاوی هسته متصل به غشاء هستند که در آن DNA در اطراف پروتئینهای هیستون پیچیده شده است. چند نوع قارچ ساختاری قابل مقایسه با پلاسمیدهای باکتریایی (حلقه های DNA) دارند. سلولهای قارچی همچنین حاوی میتوکندری و سیستم پیچیدهای از غشای داخلی، از جمله شبکه آندوپلاسمی و دستگاه گلژی هستند.
برخلاف سلولهای گیاهی، سلولهای قارچی کلروپلاست یا کلروفیل ندارند. بسیاری از قارچها رنگهای روشن ناشی از رنگدانههای سلولی دیگر، از قرمز تا سبز و سیاه را نشان میدهند. رنگدانهها در قارچها با دیواره سلولی مرتبط هستند. آنها نقش محافظتی در برابر اشعه ماوراء بنفش دارند و میتوانند سمی باشند.

لایههای سفت و سخت دیواره سلولی قارچ حاوی پلیساکاریدهای پیچیدهای به نام کیتین و گلوکان است. کیتین که در اسکلت بیرونی حشرات نیز یافت میشود، به دیواره سلولی قارچها استحکام ساختاری میبخشد. دیواره سلول را از خشک شدن و شکارچیان محافظت میکند. قارچها دارای غشای پلاسمایی مشابه سایر یوکاریوتها هستند، با این تفاوت که این ساختار توسط ارگوسترول تثبیت میشود. ارگوسترول یک مولکول استروئیدی بوده که جایگزین کلسترول موجود در غشای سلولی حیوانات میشود. اکثر اعضای پادشاهی قارچها غیر متحرک هستند.
بخش های مختلف سلول یوکاریوت
ارگانیسمهای یوکاریوتی شامل تکیاختهها، جلبکها، قارچها، گیاهان و جانوران هستند. برخی از سلولهای یوکاریوتی میکروارگانیسمهای مستقل و تکسلولی بوده، در حالی که برخی دیگر از موجودات چندسلولی هستند. سلولهای موجودات یوکاریوتی دارای چندین ویژگی متمایز هستند. مهمتر از همه، سلولهای یوکاریوتی با حضور هستهای که توسط غشای هستهای پیچیده احاطه شده است، تعریف میشوند. همچنین سلولهای یوکاریوتی با وجود اندامکهای متصل به غشاء در سیتوپلاسم مشخص میشوند.
اندامکهایی مانند میتوکندری، شبکه آندوپلاسمی (ER)، دستگاه گلژی، لیزوزومها و پراکسیزومها توسط اسکلت سلولی (شبکه داخلی که از انتقال اجزای درون سلولی پشتیبانی کرده و به حفظ شکل سلول کمک میکند)، در جای خود نگه داشته میشوند. ژنوم سلولهای یوکاریوتی بر خلاف کروموزوم منفرد دایرهای شکل که مشخصه اکثر سلولهای پروکاریوتی است، در کروموزومهای میلهای شکل متعدد بستهبندی میشود. در بخشهای قبل به طور خلاصه اجزای انواع سلولهای یوکاریوت را نام بردیم در این بخش اجزای ساختاری سلول یوکاریوتی را به طور کاملتر و دقیقتر بررسی میکنیم.
هسته
برخلاف سلولهای پروکاریوتی، که در آنها DNA آزادانه در ناحیه نوکلوئیدی قرار دارد، سلولهای یوکاریوتی دارای هسته بوده که توسط غشای هستهای احاطه شده و ژنوم DNA را در خود جای داده است. با داشتن DNA سلول، هسته در نهایت تمام فعالیتهای سلول را کنترل میکند و همچنین نقش اساسی در تولید مثل و وراثت دارد. سلولهای یوکاریوتی معمولاً DNA خود را در کروموزومهای خطی متعدد سازماندهی میکنند. DNA بسیار سازماندهی شده و متراکم شده است تا درون هسته قرار گیرد، این فرایند با پیچیدن DNA در اطراف پروتئینهایی به نام هیستون انجام میشود.

مثال هایی از یوکاریوت های چند هسته ای
اگرچه اکثر سلولهای یوکاریوتی فقط یک هسته دارند، استثناهایی وجود دارد. برای مثال، تکیاختههای جنس Paramecium معمولاً دو هسته کامل دارند. در این موجودات هسته کوچک برای تولیدمثل استفاده میشود (میکرونوکلئوس) و هسته بزرگ (ماکرونوکلئوس) متابولیسم سلولی را هدایت میکند. علاوه بر این، برخی از قارچها در طول تولیدمثل جنسی، سلولهایی با دو هسته به نام سلولهای هتروکاریوتی تشکیل میدهند. سلولهایی که هسته آنها تقسیم میشود، اما سیتوپلاسم آنها تقسیم نمیشود، کوئنوسیت نامیده میشوند.
ساختار هسته
هسته توسط غشای هستهای پیچیده که اغلب پوشش هستهای نامیده شده، محدود میشود. این غشا از یک ساختار دولایه لیپیدی مجزا تشکیل شده است که با یکدیگر به هم پیوستهاند. با وجود این اتصالات بین غشاهای داخلی و خارجی، هر غشا حاوی لیپیدها و پروتئینهای منحصر به فردی در سطوح داخلی و خارجی خود است. پوشش هستهای حاوی منافذ هستهای است که کمپلکسهای پروتئینی بزرگ اطراف این منافظ قرار گرفته و حرکت مواد به داخل و خارج از هسته را کنترل میکنند. شکل کلی هسته توسط لامینای هستهای تعیین میشود، شبکهای از رشتههای میانی که درست در داخل غشای پوشش هستهای یافت میشود. در خارج از هسته، رشتههای میانی اضافی شبکه شلتری را تشکیل میدهند و برای لنگر انداختن هسته در موقعیت درون سلول عمل میکنند.
هستک
هستک یک ناحیه متراکم درون هسته است که در آن بیوسنتز RNA ریبوزومی (rRNA) اتفاق میافتد. علاوه بر این، هستک همچنین محلی است که در آن مونتاژ ریبوزومها شروع میشود. برخی از کروموزومها دارای بخشهایی از DNA هستند که RNA ریبوزومی را کد میکند. مجتمعهای پیشریبوزومی از rRNA و پروتئینها به زیرواحدهای ریبوزومی در هستک جمع میشوند. آنها سپس به سیتوپلاسم منتقل میشوند، جایی که مونتاژ ریبوزوم تکمیل میشود.

کروماتین و کروموزوم
بین هستک و پوشش هسته کروماتین قرار دارد. برای درک کروماتین، ابتدا کروموزومها را در نظر بگیرید. کروموزومها ساختارهایی در هسته بوده که از DNA، ماده ارثی و حامل اطلاعات (ژن) مورد نیاز ساخته شدهاند. کروموزومهای یوکاریوتی معمولاً خطی هستند و سلولهای یوکاریوتی چندین کروموزوم مجزا دارند. بسیاری از سلولهای یوکاریوتی حاوی دو نسخه از هر کروموزوم و بنابراین دیپلوئید هستند. طول کروموزوم بسیار بیشتر از طول سلول است، بنابراین کروموزوم باید در یک فضای بسیار کوچک بستهبندی شود تا در داخل سلول قرار گیرد.
به عنوان مثال، طول مجموع تمام DNA ژنوم انسان اگر به طور کامل کشیده شود تقریباً 2 متر خواهد بود و برخی از ژنومهای یوکاریوتی چندین برابر بزرگتر از ژنوم انسان هستند. هر گونه یوکاریوتی تعداد مشخصی کروموزوم در هسته سلولهای بدن خود دارد. به عنوان مثال، در انسان، عدد کروموزوم 46 است، در حالی که در مگس میوه، 8 عدد است. کروموزومها تنها زمانی قابل مشاهده و تشخیص از یکدیگر هستند که سلول در حال آماده شدن برای تقسیم است.
هنگامی که سلول در مراحل رشد و نگهداری از چرخه زندگی خود است، پروتئینها به کروموزومها متصل شده و شبیه به دستهای از رشتههای درهم ریخته و درهم پیچیده میشوند. به این کمپلکسهای کروموزوم-پروتئین باز نشده کروماتین میگویند.
ریبوزوم
ریبوزومهای سیتوپلاسمی در سلولهای یوکاریوتی ریبوزومهای 80S هستند که از زیرواحد کوچک 40S و زیرواحد بزرگ 60S تشکیل شدهاند. از نظر اندازه و ترکیب، این تفاوت ساختاری آنها را از ریبوزومهای سلولهای پروکاریوتی متمایز میکند. دو نوع ریبوزوم یوکاریوتی سیتوپلاسمی بر اساس مکان آنها در سلول تعریف میشوند: ریبوزومهای آزاد و ریبوزومهای متصل به غشاء. ریبوزومهای آزاد در سیتوپلاسم شناور یافت میشوند و برای سنتز پروتئینهای محلول در آب عمل میکنند. ریبوزومهای متصل به غشاء در سمت سیتوپلاسمی شبکه آندوپلاسمی زبر یافت میشوند و پروتئینهایی را برای وارد کردن به غشای سلولی ایجاد کرده یا پروتئینهایی میسازند که قرار است از سلول خارج شوند.
در مقابل، ریبوزومهای موجود در اندامکهای یوکاریوتی مانند میتوکندری یا کلروپلاست، ریبوزومهای 70S دارند که به اندازه ریبوزومهای پروکاریوتی است. تفاوت بین ریبوزومهای یوکاریوتی و پروکاریوتی از نظر بالینی مهم است زیرا داروهای آنتیبیوتیک خاصی برای هدف قرار دادن یکی یا دیگری طراحی شدهاند. به عنوان مثال، سیکلوهگزیماید عملکرد ریبوزوم یوکاریوتی را هدف قرار میدهد، در حالی که کلرامفنیکل ریبوزومهای پروکاریوتی را هدف قرار میدهد. از آنجایی که سلولهای انسانی یوکاریوتی هستند، معمولاً توسط آنتیبیوتیکهایی که ریبوزومهای پروکاریوتی موجود در باکتریها را از بین میبرند، آسیبی نمیبینند. با این حال، گاهی اوقات ممکن است عوارض جانبی منفی رخ دهد زیرا میتوکندری موجود در سلولهای انسانی حاوی ریبوزومهای پروکاریوتی است.

سیستم غشایی داخلی
سیستم غشایی داخلی که منحصر به سلولهای یوکاریوتی است، مجموعهای از لولههای غشایی، کیسهها و دیسکهای مسطح بوده که بسیاری از اجزای سلولی را سنتز میکنند و مواد را در داخل سلول حرکت میدهند. سلولهای یوکاریوتی به دلیل اندازه سلول بزرگتر خود به این سیستم برای انتقال موادی نیاز دارند که نمیتوانند به تنهایی از طریق انتشار پراکنده شوند. سیستم غشایی داخلی شامل چندین اندامک و اتصالات بین آنها، از جمله شبکه آندوپلاسمی، دستگاه گلژی، لیزوزومها و وزیکولها است. در ادامه بخشهای مختلف سیستم غشایی داخلی را در یوکاریوتها بررسی میکنیم.
شبکه آندوپلاسمی
شبکه آندوپلاسمی (ER) یک آرایه به هم پیوسته از لولهها و سیسترناها (کیسههای مسطح) با یک لایه لیپیدی منفرد است. فضاهای داخل سیسترنا لومن شبکه اندوپلاسمی نامیده میشود. دو نوع شبکه اندوپلاسمی وجود دارد، «شبکه آندوپلاسمی خشن/زبر» (RER) و «شبکه آندوپلاسمی صاف» (SER). این دو نوع مختلف ER مکانهایی برای سنتز انواع مختلف مولکولها هستند. شبکه اندوپلاسمی زبر با ریبوزومهای متصل به سمت سیتوپلاسمی غشاء پوشانده شده است. این ریبوزومها پروتئینهایی را برای غشای پلاسمایی تولید میکنند. پس از سنتز، این پروتئینها به غشای RER وارد میشوند.
کیسههای کوچک شبکه اندوپلاسمی زبر حاوی این پروتئینهای تازه سنتز شده، جوانه میزنند و برای پردازش بیشتر، مستقیماً به غشای پلاسمایی، غشای اندامک دیگری یا دستگاه گلژی و سپس خارج از سلول میروند. شبکه اندوپلاسمی صاف ریبوزوم ندارد و بنابراین «صاف» به نظر میرسد. این بخش در بیوسنتز لیپیدها، سنتز و تجزیه کربوهیدراتها، سمزدایی داروها و سموم نقش و ذخیره یونهای کلسیم نقش دارد.

وزیکول ها و واکوئل ها
وزیکولها و واکوئلها کرههای غشایی تکلیپیدی، دولایهای با فضای داخلی توخالی هستند که در ذخیرهسازی و حمل و نقل عمل میکنند. به غیر از این واقعیت که واکوئلها تا حدودی بزرگتر از وزیکولها هستند، یک تمایز بسیار ظریف بین آنها وجود دارد: غشاهای وزیکولها میتوانند با غشای پلاسمایی یا سایر سیستمهای غشایی درون سلول ترکیب شوند. علاوه بر این، برخی از عوامل مانند آنزیمهای درون واکوئلهای گیاهی، ماکرومولکولها را تجزیه میکنند. غشای واکوئل با غشای سایر اجزای سلولی ترکیب نمیشود و همین امر آنها را به واحدهای ذخیرهسازی عالی تبدیل میکند.
وزیکولهایی که از ER خارج میشوند با پروتئینها، کربوهیدراتها و سایر کالاها یا ضایعات با دستگاه گلژی بر روی صفحه گیرنده یا سیس آن ترکیب میشوند. پروتئینها در دستگاه گلژی پردازش میشوند و سپس وزیکولهای حمل و نقل اضافی حاوی پروتئینها و لیپیدهای اصلاح شده از دستگاه گلژی روی بخش خروجی یا ترانس آن جدا میشوند. این وزیکولهای خروجی به غشای پلاسمایی (از طریق اگزوسیتوز) یا غشای سایر اندامکها حرکت میکنند و با آن ترکیب میشوند.
دستگاه گلژی
دستگاه گلژی در سال 1898 توسط دانشمند ایتالیایی کامیلو گلژی (1843-1926) در سیستم غشای داخلی یوکاریوتها کشف شد. این دانشمند یک تکنیک رنگآمیزی جدید را توسعه داد که ساختارهای غشایی انباشته شده را در سلولهای پلاسمودیوم (عامل ایجاد مالاریا) نشان داد. دستگاه گلژی از مجموعهای از دیسکهای غشایی به نام دیکتوزومها تشکیل شده است که هر کدام دارای یک لایه لیپیدی منفرد هستند که در کنار هم قرار گرفتهاند. اصلاح، مرتبسازی، برچسبگذاری، بستهبندی و توزیع لیپیدها و پروتئینها در دستگاه گلژی صورت میگیرد.

آنزیمهای موجود در دستگاه گلژی، لیپیدها و پروتئینهای منتقل شده از شبکه اندوپلاسمی به گلژی را اصلاح میکنند، اغلب اجزای کربوهیدراتی را به آنها اضافه کرده و گلیکولیپیدها، گلیکوپروتئینها یا پروتئوگلیکانها را تولید میکنند. گلیکولیپیدها و گلیکوپروتئینها اغلب به غشای پلاسمایی وارد میشوند و برای تشخیص سیگنال توسط سلولهای دیگر یا ذرات عفونی مهم هستند. انواع مختلف سلولها را میتوان با ساختار و آرایش گلیکولیپیدها و گلیکوپروتئینهای موجود در غشای پلاسمایی آنها از یکدیگر متمایز کرد. این گلیکولیپیدها و گلیکوپروتئینها معمولاً به عنوان گیرندههای سطح سلولی نیز عمل میکنند.
لیزوزوم ها
در دهه 1960، دانشمند بلژیکی کریستین دوو لیزوزومها، اندامکهای متصل به غشای سیستم غشایی داخلی را کشف کرد که حاوی آنزیمهای گوارشی هستند. انواع خاصی از سلولهای یوکاریوتی از لیزوزومها برای تجزیه ذرات مختلف مانند غذا، اندامکهای آسیبدیده یا بقایای سلولی، میکروارگانیسمها یا کمپلکسهای ایمنی استفاده میکنند. تقسیمبندی آنزیمهای گوارشی در لیزوزوم به سلول اجازه میدهد تا ماده را بدون آسیب رساندن به اجزای سیتوپلاسمی سلول هضم کند.
پراکسی زوم ها
کریستین دی دوو همچنین برای کشف پراکسیزومها، اندامکهای متصل به غشاء که بخشی از سیستم غشایی داخلی نیستند، شناخته شده است. پراکسیزومها به طور مستقل در سیتوپلاسم از سنتز پروتئینهای پروکسین توسط ریبوزومهای آزاد و ادغام این پروتئینهای پروکسین به پراکسیزومهای موجود تشکیل میشوند. سپس پراکسیزومهای در حال رشد با فرآیندی مشابه شکافت دوتایی تقسیم میشوند.

پراکسیزومها اولین بار به دلیل توانایی آنها در تولید پراکسید هیدروژن، یک مولکول بسیار واکنشپذیر که به تجزیه مولکولهایی مانند اسیداوریک، اسیدهای آمینه و اسیدهای چرب کمک میکند، نامگذاری شدند. پراکسیزومها همچنین دارای آنزیم کاتالاز هستند که میتواند پراکسید هیدروژن را تجزیه کند. همراه با شبکه اندوپلاسمی صاف، پراکسیزومها در بیوسنتز لیپیدها نیز نقش دارند. مانند لیزوزومها، تقسیم این مولکولهای تخریبکننده در یک اندامک به محافظت از محتویات سیتوپلاسمی در برابر آسیبهای ناخواسته کمک میکند.
پراکسیزومهای موجودات خاص برای برآوردن نیازهای عملکردی خاص آنها تخصصی شده است. به عنوان مثال، گلیاکسیزومها پراکسیزومهای اصلاحشده مخمرها و سلولهای گیاهی هستند که چندین عملکرد متابولیکی از جمله تولید مولکولهای قند را انجام میدهند. به طور مشابه، گلیکوزومها پراکسیزومهای اصلاح شده ساخته شده توسط تریپانوزومهای خاص، تکیاختههای بیماریزا هستند که باعث بیماری شاگاس و بیماری خواب آفریقایی میشوند.
اسکلت سلولی
سلولهای یوکاریوتی دارای اسکلت سلولی داخلی هستند که از ریزرشتهها، رشتههای میانی و میکروتوبولها ساخته شده است. این ماتریس از رشتهها و لولهها پشتیبانی ساختاری و همچنین شبکهای را فراهم میکند که مواد را میتوان از طریق آن در داخل سلول حمل کرد و اندامکها را میتوان بر روی آن لنگر انداخت. به عنوان مثال، فرآیند اگزوسیتوز شامل حرکت وزیکول از طریق شبکه اسکلت سلولی به غشای پلاسمایی است، جایی که میتواند محتویات خود را آزاد کند. ریزرشتهها از دو رشته اکتین در هم تنیده تشکیل شدهاند که هر کدام از مونومرهای اکتین تشکیل شدهاند که کابلهای رشتهای به قطر ۶ نانومتر را تشکیل میدهند.

رشتههای اکتین با پروتئینهای حرکتی مانند میوزین کار میکنند تا انقباض ماهیچهای در حیوانات یا حرکت آمیبوئید برخی از میکروبهای یوکاریوتی را تحت تأثیر قرار دهند. در موجودات آمبوئید، اکتین را میتوان به دو شکل یافت: یک فرم سفتتر، پلیمریزه شده، به شکل ژل و یک شکل محلول سیالتر و غیر پلیمریزه. اکتین به شکل ژل باعث ایجاد ثبات در اکتوپلاسم میشود، ناحیه ژلمانند سیتوپلاسم درست در داخل غشای پلاسمایی تکیاختههای آمبوئید. گسترش موقت غشای سیتوپلاسمی به نام «پاهای کاذب» (Pseudopodia) از طریق جریان رو به جلو رشتههای اکتین محلول به داخل پای کاذب، به دنبال چرخه ژل-سلول رشتههای اکتین تولید میشوند که منجر به تحرک سلولی خواهد شد.
هنگامی که سیتوپلاسم به سمت بیرون گسترش مییابد و پای کاذب را تشکیل میدهد، سیتوپلاسم باقیمانده برای پیوستن به لبه جلویی جریان پیدا کرده و در نتیجه حرکت رو به جلو ایجاد میکند. فراتر از حرکت آمیبوئید، ریزرشتهها همچنین در انواع فرآیندهای دیگر در سلولهای یوکاریوتی، از جمله جریان سیتوپلاسمی (حرکت یا گردش سیتوپلاسم در داخل سلول)، تشکیل شیارهای برش در طول تقسیم سلولی و حرکت ماهیچهها در حیوانات دخیل هستند. این عملکردها نتیجه ماهیت دینامیکی ریزرشتهها است که میتوانند نسبتاً به راحتی در پاسخ به سیگنالهای سلولی پلیمریزه و دپلیمریزه شوند و برهمکنش آنها با موتورهای مولکولی در انواع مختلف سلولهای یوکاریوتی رخ میدهد.
فیلامانهای حد واسط گروه متنوعی از رشتههای اسکلت سلولی هستند که به عنوان کابل در داخل سلول عمل میکنند. آنها متوسط نامیده میشوند زیرا قطر 10 نانومتری آنها ضخیمتر از قطر اکتین اما نازکتر از قطر ریز لولهها است. آنها از چندین رشته از زیرواحدهای پلیمریزه شده تشکیل شدهاند که به نوبه خود از طیف گستردهای از مونومرها ساخته شدهاند. رشتههای میانی تمایل بیشتری به ماندگاری در سلول دارند و موقعیت هسته را حفظ میکنند. آنها همچنین لایه هستهای (پوشش یا لایه) را درست در داخل پوشش هسته تشکیل میدهند.

علاوه بر این، رشتههای میانی در پیوند دادن سلولها به یکدیگر در بافتهای حیوانی نقش دارند. پروتئین رشته میانی دسمین در دسموزومها یافت میشود، ساختارهای پروتئینی که سلولهای عضلانی را به هم میپیوندند و به آنها در مقاومت در برابر نیروهای فیزیکی خارجی کمک میکنند. پروتئین فیلامنت میانی کراتین یک پروتئین ساختاری است که در مو، پوست و ناخنها یافت میشود.
میکروتوبول ها نوع سوم فیبر اسکلت سلولی هستند که از دایمرهای توبولین (α توبولین و β توبولین) تشکیل شده است. این لولههای توخالی به قطر 23 نانومتر را تشکیل میدهند که به عنوان تیرهای درون اسکلت سلولی استفاده میشوند. مانند میکرو فیلامنتها، میکروتوبولها نیز پویا هستند و قابلیت جمع شدن و جداسازی سریع را دارند. میکروتوبولها همچنین با پروتئینهای حرکتی (مانند داینئین و کاینزین) کار میکنند تا اندامکها و وزیکولها را در داخل سیتوپلاسم حرکت دهند. میکروتوبولها نیز در تقسیم سلولی نقش دارند و دوک میتوزی را تشکیل میدهند که برای جداسازی کروموزومها در طول میتوز و میوز عمل میکند. دوک میتوزی توسط دو سانتروزوم، که اساساً مراکز سازماندهی میکروتوبول هستند، در انتهای مخالف سلول تولید میشود.

میتوکندری
میتوکندری اغلب به عنوان نیروگاه یا کارخانههای انرژی سلول نامیده میشوند زیرا آنها مسئول ساخت آدنوزین تریفسفات (ATP)، مولکول اصلی حامل انرژی سلول هستند. ATP نشاندهنده انرژی ذخیره شده کوتاهمدت سلول است. تنفس سلولی فرآیند ساخت ATP با استفاده از انرژی شیمیایی موجود در گلوکز و سایر مواد مغذی است. در میتوکندری، این فرآیند از اکسیژن استفاده میکند و دیاکسیدکربن را به عنوان یک محصول زائد تولید میکند. در واقع، دیاکسیدکربنی که با هر نفس بازدم میکنید از واکنشهای سلولی ناشی میشود که دیاکسیدکربن را به عنوان یک محصول جانبی تولید میکنند. اصطلاح میتوکندری اولین بار توسط میکروبیولوژیست آلمانی کارل بندا در سال 1898 ابداع شد و بعداً توسط اتوواربورگ در سال 1913 با فرآیند تنفس مرتبط شد.

هر میتوکندری دارای دو غشای لیپیدی است. غشای خارجی بقایای ساختارهای غشایی سلول میزبان اصلی است. غشای داخلی از غشای پلاسمایی باکتری گرفته شده است. زنجیره انتقال الکترون برای تنفس هوازی از پروتئینهای اینتگرال تعبیه شده در غشای داخلی استفاده میکند. ماتریکس میتوکندری، مربوط به محل سیتوپلاسم باکتری اصلی، محل فعلی بسیاری از آنزیمهای متابولیک است. همچنین حاوی DNA میتوکندری و ریبوزوم 70S است. تورفتگیهای غشای داخلی، به نام «کریستا» (Cristae)، برای افزایش سطح برای مکان واکنشهای بیوشیمیایی تکامل یافته است. الگوهای تاشوی کریستاها در میان انواع مختلف سلولهای یوکاریوتی متفاوت است و برای تشخیص ارگانیسمهای یوکاریوتی مختلف از یکدیگر استفاده میشود.
کلروپلاست
سلولهای گیاهی و سلولهای جلبکی حاوی کلروپلاست هستند، اندامکهایی که فتوسنتز در آنها اتفاق میافتد. فتوسنتز مجموعهای از واکنشها است که از دیاکسیدکربن، آب و انرژی نور برای تولید گلوکز و اکسیژن استفاده میکند. تمام کلروپلاستها حداقل دارای سه سیستم غشایی هستند: غشای بیرونی، غشای داخلی و سیستم غشایی تیلاکوئیدی. در داخل غشاهای بیرونی و داخلی، استرومای کلروپلاست وجود دارد، مایعی ژلمانند که بیشتر حجم کلروپلاست را تشکیل میدهد و سیستم تیلاکوئیدی در آن شناور است.
سیستم تیلاکوئید مجموعهای بسیار پویا از کیسههای غشایی چینخورده است. جایی است که رنگدانه سبز فتوسنتزی کلروفیل یافت میشود و واکنشهای نوری فتوسنتز رخ میدهد. در بیشتر کلروپلاستهای گیاهی، تیلاکوئیدها در پشتههایی به نام گرانا (مفرد: گرانوم) مرتب شدهاند، در حالی که در برخی از کلروپلاستهای جلبکی، تیلاکوئیدها به صورت آزاد شناور هستند. سایر اندامکهای مشابه میتوکندری در انواع دیگر یوکاریوتها به وجود آمدهاند، اما نقش آنها متفاوت است. هیدروژنوزومها در برخی از یوکاریوتهای بیهوازی یافت میشوند و به عنوان محل تولید هیدروژن بیهوازی عمل میکنند.
هیدروژنوزومها معمولاً فاقد DNA و ریبوزوم خاص خود هستند. کینتوپلاستها گونهای از میتوکندری هستند که در برخی از پاتوژنهای یوکاریوتی یافت میشوند. در این موجودات، هر سلول دارای میتوکندری منفرد، طویل و شاخهدار است که در آن DNA کینتوپلاست، که به صورت تکههای دایرهای متعدد از DNA سازماندهی شده و در قطب سلول متمرکز شده است.

غشای سلولی
ساختار غشای پلاسمایی اکثر انواع سلولهای باکتریایی و یوکاریوتی به صورت دولایه بوده که عمدتاً از فسفولیپیدهایی تشکیل شده است که با پیوندهای استری و پروتئینها تشکیل شده است. پروتئینهای روی سطح سلول برای عملکردهای مختلف مهم هستند. برخلاف غشای پروکاریوتی، غشاهای یوکاریوتی حاوی استرولهایی از جمله کلسترول هستند که سیالیت غشاء را تغییر میدهند. علاوه بر این، بسیاری از سلولهای یوکاریوتی حاوی برخی لیپیدهای تخصصی، از جمله اسفنگولیپیدها هستند که تصور میشود در حفظ ثبات غشاء و همچنین درگیر شدن در مسیرهای انتقال سیگنال و ارتباط سلول به سلول نقش دارند.
دیواره سلولی
علاوه بر غشای پلاسمایی، برخی از سلولهای یوکاریوتی دارای دیواره سلولی هستند. سلولهای قارچها، جلبکها، گیاهان و حتی برخی از آغازیان دارای دیوارههای سلولی هستند. بسته به نوع سلول یوکاریوتی، دیوارههای سلولی میتوانند از طیف وسیعی از مواد، از جمله سلولز (در قارچها و گیاهان)، سیلیس، کربنات کلسیم، آگار و کاراگینان (در آغازیان و جلبکها) یا کیتین (در قارچ) ساخته شوند. به طور کلی، تمام دیوارههای سلولی پایداری ساختاری سلول و محافظت در برابر تنشهای محیطی مانند خشک شدن، تغییرات فشار اسمزی و آسیبهای تروماتیک را فراهم میکنند.
ماتریکس خارج سلولی
سلولهای جانوران و برخی تکیاختهها دیواره سلولی ندارند تا به حفظ شکل و ایجاد ثبات ساختاری کمک کند. در عوض، بسیاری از این نوع سلولهای یوکاریوتی ماتریکس خارج سلولی برای این منظور تولید میکنند. آنها توده چسبندهای از کربوهیدراتها و پروتئینها را در فضای بین سلولهای مجاور ترشح میکنند. برخی از اجزای پروتئین در غشای پایه جمع میشوند که اجزای ماتریکس خارج سلولی باقیمانده به آن میچسبند.

پروتئوگلیکانها معمولاً توده حجیم ماتریکس خارج سلولی را تشکیل میدهند در حالی که پروتئینهای فیبری مانند کلاژن استحکام میبخشند. پروتئوگلیکانها و کلاژن هر دو به پروتئینهای فیبرونکتین متصل هستند که به نوبه خود به پروتئینهای اینتگرین متصل میشوند. این پروتئینهای اینتگرین با پروتئینهای گذرنده در غشای پلاسمایی سلولهای یوکاریوتی که فاقد دیواره سلولی هستند، تعامل دارند. این مسئله باعث میشود که آنها هر دو مشابه و متفاوت با گلیکوکالیکس موجود در سلولهای پروکاریوت باشند.
در سلولهای حیوانی، ماتریکس خارج سلولی به سلولهای درون بافت اجازه میدهد تا در برابر استرسهای خارجی مقاومت کنند و سیگنالها را از بیرون سلول به داخل منتقل میکنند. مقدار ماتریکس خارج سلولی در انواع مختلف بافت همبند بسیار گسترده است و تغییرات در ماتریکس خارج سلولی میتواند به انواع مختلف بافتها خواص متمایز آنها را بدهد. علاوه بر این، ماتریکس خارج سلولی سلول میزبان اغلب محلی است که پاتوژنهای میکروبی خود را برای ایجاد عفونت میچسبانند. به عنوان مثال، استرپتوکوک پیوژنز، باکتری که باعث گلودرد استرپتوکوکی و عفونتهای مختلف دیگر میشود، به فیبرونکتین در ماتریکس خارج سلولی سلولهای پوشاننده اوروفارنکس (ناحیه بالایی گلو) متصل میشود.
تاژک و مژک در یوکاریوت چگونه است؟
برخی از سلولهای یوکاریوتی از تاژکها برای حرکت استفاده میکنند. با این حال، تاژکهای یوکاریوتی از نظر ساختاری از آنهایی که در سلولهای پروکاریوتی یافت میشوند متمایز هستند. در حالی که تاژک پروکاریوتی یکساختار سفت و چرخان است، تاژک یوکاریوتی بیشتر شبیه یک شلاق انعطافپذیر است که از 9 جفت ریزلوله موازی تشکیل شده که یک جفت میکروتوبول مرکزی را احاطه کرده است. این آرایش به عنوان آرایه 9+2 نامیده میشود. میکروتوبولهای موازی از پروتئینهای حرکتی داینئین برای حرکت نسبت به یکدیگر استفاده میکنند و باعث خم شدن تاژک میشوند. مژک ساختار خارجی مشابهی است که در برخی از سلولهای یوکاریوتی یافت میشود.

مژکها که منحصر به یوکاریوتها هستند، کوتاهتر از تاژکها هستند و اغلب تمام سطح سلول را میپوشانند. با این حال، آنها از نظر ساختاری شبیه تاژکها (آرایه 9+2 از میکروتوبولها) هستند و از مکانیسم یکسانی برای حرکت استفاده میکنند. ساختاری به نام جسم بازال در پایه هر مژک و تاژک یافت میشود. جسم بازال که به اتصال مژک یا تاژک به سلول کمک میکند از مجموعهای از میکرولولههای سهگانه شبیه به یک سانتریول تشکیل شده است اما در غشای پلاسمایی جاسازی شده است.
به دلیل طول کوتاهتر، مژکها از حرکت سریع، انعطافپذیر و موجدار استفاده میکنند. علاوه بر تحرک، مژکها ممکن است عملکردهای دیگری مانند جارو کردن ذرات از گذشته یا درون سلولها را داشته باشند. به عنوان مثال، تکیاختههای مژکدار از حرکت شبیه جارو کردن مژکها برای انتقال ذرات غذا به داخل قسمتهای دهان خود استفاده میکنند، و سلولهای مژکدار در دستگاه تنفسی پستانداران به طور همزمان برای جارو کردن مخاط و باقیماندهها به سمت بالا و خارج از ریهها حرکت میکنند.
طبقه بندی یوکاریوت ها
یوکاریوتها حوزهای از حیات را نشان میدهند، اما در این حوزه فرمانروهای متعددی وجود دارد. رایجترین طبقهبندی چهار فرمانرو را در این حوزه ایجاد میکند: «گیاهان، جانوران، قارچها و آغازیان» (Protista، Fungi، Plantae و Animalia).این حوزهها بیشتر به دستههای کوچکتر زیادی تقسیم میشوند: فیلا (مفرد: شاخه)، طبقات، راستهها، خانوادهها، جنسها و گونهها. شما میتوانید تصور کنید که حتی در یک حوزه، بسیاری از ارگانیسمهای مختلف وجود دارند که ممکن است خیلی شبیه به هم نباشند یا اعمال متفاوتی داشته باشند. در ادامه انواع یوکاریوتها را بررسی میکنیم.
آغازیان چیست؟
فرمانرو «آغازیان» (Protista) از پروتیستهای یوکاریوتی تشکیل شده است. اعضای این پادشاهی بسیار متنوع معمولاً تکسلولی هستند و از نظر ساختار پیچیدهتر از سایر یوکاریوتها هستند. در یک مفهوم سطحی، این موجودات اغلب بر اساس شباهت آنها به گروههای دیگر یوکاریوتها: حیوانات، گیاهان و قارچها توصیف میشوند. آغازیان شباهتهای زیادی با هم ندارند، اما به دلیل اینکه در هیچ یک از فرمانروهای دیگر نمیگنجند، باهم در یک گروه قرار میگیرند. برخی از پروتیستها قادر به فتوسنتز هستند. برخی در روابط متقابل با دیگر آغازیان زندگی میکنند.

برخی تکسلولی هستند. برخی از آنها چندسلولی بوده یا کلونی تشکیل میدهند. انواعی از آنها همانطور که در شکل بالا نشان داده شده است میکروسکوپی هستند. بعضی از آغازیان دارای خاصیت بیولومنسنت هستند و برخی مسئول تعدادی از بیماریهایی هستند که در گیاهان و حیوانات رخ میدهد. آغازیان در محیطهای آبی، زیستگاههای خشکی مرطوب و حتی در داخل سایر یوکاریوتها زندگی میکنند.
ویژگیهای آغازیان
پروتیستها در زیر دامنه یوکاریا زندگی میکنند و بنابراین به عنوان یوکاریوتها طبقهبندی میشوند. موجودات یوکاریوتی از پروکاریوتها متمایز میشوند زیرا دارای هستهای هستند که توسط غشاء احاطه شده است. علاوه بر هسته، آغازیان اندامکهای اضافی در سیتوپلاسم خود دارند. شبکه آندوپلاسمی و مجتمعهای گلژی برای سنتز پروتئینها و اگزوسیتوز مولکولهای سلولی مهم هستند. بسیاری از پروتیستها دارای لیزوزوم هستند که به هضم مواد آلی بلعیده شده کمک میکند. اندامکهای خاصی ممکن است در برخی از سلولهای پروتیست یافت شوند و در برخی دیگر نه.
پروتیستهایی که ویژگیهای مشترک با سلولهای حیوانی دارند، میتوکندری نیز دارند که انرژی سلول را تامین میکند. آغازیانی که شبیه سلولهای گیاهی هستند دارای دیواره سلولی و کلروپلاست هستند. کلروپلاستها فتوسنتز را در این سلولها ممکن میسازند. در ادامه ویژگیهای اصلی آغازیان را معرفی میکنیم.
حرکت در آغازیان
در حالی که برخی از پروتیستها غیرمتحرک هستند، برخی دیگر حرکت را از طریق روشهای مختلف نشان میدهند. برخی از آغازیان تاژک یا مژک دارند. این اندامکها برآمدگیهایی هستند که از گروهبندیهای تخصصی ریزلولهها تشکیل شدهاند که برای به حرکت درآوردن انواع آغازیان در محیط مرطوب خود حرکت میکنند. سایر پروتیستها با استفاده از امتداد موقت سیتوپلاسم خود به نام پای کاذب حرکت میکنند. این گسترشها همچنین برای اینکه به پروتیست اجازه میدهند سایر ارگانیسمهایی را که از آنها تغذیه میکنند، بگیرد، ارزشمند هستند.
نحوه تغذیه آغازیان
آغازیان روشهای مختلفی را برای کسب غذا نشان میدهند. برخی از آنها اتوتروفهای فتوسنتزی بوده، به این معنی که آنها خود غذاساز و قادر به استفاده از نور خورشید برای تولید کربوهیدرات جهت تغذیه هستند. دیگر پروتیستها هتروتروفها هستند که از موجودات دیگر تغذیه میکنند. این امر توسط فاگوسیتوز انجام میشود، فرآیندی که در آن ذرات در داخل فرورفته و هضم میشوند. با این حال، سایر آغازیان عمدتاً با جذب مواد مغذی از محیط خود تغذیه میکنند. برخی از پروتیستها ممکن است هر دو شکل فتوسنتزی و هتروتروف جذب مواد مغذی را نشان دهند.
تولید مثل آغازیان
متداولترین روش تولید مثل که در پروتیستها نمایش داده میشود، تولیدمثل غیرجنسی است. تولیدمثل جنسی امکانپذیر است، اما معمولاً فقط در زمان استرس رخ میدهد. برخی از پروتیستها به صورت غیرجنسی با شکافت دوتایی یا شکافت چندگانه تولیدمثل میکنند. برخی دیگر با جوانهزدن یا از طریق تشکیل هاگ به صورت غیرجنسی تولید مثل میکنند. در تولید مثل جنسی، گامتها توسط میوز تولید میشوند و در زمان لقاح برای تولید افراد جدید متحد میشوند. سایر پروتیستها، مانند جلبکها، نوعی تناوب نسلها را نشان میدهند که در آن در چرخه زندگی خود بین مراحل هاپلوئید و دیپلوئید متناوب میشوند.
جانوران
قلمرو حیوانات از جانوران یوکاریوتی چندسلولی مختلف تشکیل شده است. این یکی از فرمانروها در میان گروههای پنجگانه موجودات است. چند ویژگی وجود دارد که برای موجودات مختلف این گروه مهم است. این اجزا شامل تقارن بدن، ماهیت سلوم، آرایش سلولها، نوتوکورد، الگوهای چارچوبهای گردش خون، گوارش و تولید مثل، تقسیمبندی و آرایش سلولها در لایههای زاینده است. هر موجودی در قلمرو حیوانات چندسلولی است. با این حال، آنها نمونه مشابهی از سازماندهی سلولی را نشان میدهند. نمونههایی از سازماندهی سلولی موجود در حیوانات عبارتند از:

- سطح سازماندهی سلولی: سلولهای حیوانات با این نوع سازماندهی سلولی در تودههای سلولی شل دستهبندی میشوند. سازماندهی اسفنجی نمونه خوبی برای این موضوع است.
- سطح سازماندهی بافتی: سلولهای حیوانی تقسیم بافتها را در میان خود نشان میدهد. سلولهایی که ظرفیت یکسانی دارند به عنوان بافت سازماندهی میشوند.
- سطح سازماندهی اندام: بافتهای یک گروه حیوانی خاص که عملکرد مشابهی را انجام میدهند با هم گروهبندی میشوند تا یک اندام ایجاد کنند. هر اندام هدف مشخصی دارد. اندامهای داخلی بدن انسان مانند معده یا روده مثالهایی از سازماندهی اندام هستند.
- سطح سازماندهی همکاری اندامها: سطح سازماندهی سیستم اندامی در حیوانات یافت شده است که در آن اندامها برای ایجاد سیستمهای عملکردی جفت شدهاند، هر سیستم با عملکرد فیزیولوژیکی خاصی مرتبط است. مانند سیستم گوارش در انسان که شامل چندین اندام مختلف و همکاری بین آنها است.
مهره داران
همه موجوداتی هستند که به زیر شاخه Vertebrata تعلق دارند. آنها اعضای گروه «مهرهداران» (Chordata) هستند و دارای یک ستون فقرات (مهره) هستند که محل قرارگیری نخاع است. علاوه بر این، آنها یک سیستم اسکلتی داخلی دارند که انواع ماهیچهها به آن متصل میشوند. در ادامه انواع مهرهداران را بررسی کردهایم.
- پستانداران: مادهها در این جانوران دارای غدد پستانی بوده که برای تغذیه به بچهها شیر تولید میکنند، دارای یک نئوکورتکس (بخشی از سیستم عصبی)، خز یا مو و سه استخوان گوش میانی (مشخصه پستانداران) هستند. این ویژگیها آنها را از خزندگان و پرندگان متمایز میکند.
- خزندگان: خزندگان (خونسرد) حیواناتی چهار پا هستند و با وجود دم و فلسهای پوستی مشخص میشوند. برای اکثر گونهها، تخمها به صورت داخلی بارور میشوند که منجر به رشد مستقیم ارگانیسم میشود. نمونههای خزندگان شامل لاکپشتها، مارها و تمساحها در میان دیگران است.
- دوزیستان: دوزیستان شامل حیوانات خونسردی هستند که بخشی از زندگی خود را در آب میگذرانند. در حالی که آنها میتوانند اکسیژن را از طریق پوست خود در یک محیط مرطوب جذب کنند، برخی از گونهها (یا در برخی از مراحل زندگی خود) دارای آبششهایی هستند که برای تنفس در آب استفاده میشود.
- ماهیها: شامل چند گروه است گروه اول ماهیهای واقعی هستند و اغلب به عنوان ماهی استخوانی شناخته میشوند. به این ترتیب، آنها با بافت استخوانی (به جای غضروف) مشخص میشوند. برخی از ویژگیهای دیگر مرتبط با این گروه شامل دندانهای جوشخورده، بالههای لوبدار و جمجمه است. اعضای این گروه شامل دلقکماهی و ماهی پرتویی هستند. ماهیهای غضروفی دارای اسکلت غضروفی بوده و به همین دلیل به ماهیهای غضروفی معروف هستند. در حالی که گونههای کمی در محیطهای آب شیرین شناسایی شدهاند، اکثر گونهها در اقیانوس یافت میشوند.
- پرندگان: پرندگان، شامل بیش از 10400 گونه زنده هستند. پرهای آنها باعث تمایز آنها از سایر حیوانات میشود. پرندگان نیز مانند پستانداران مهرهداران خونگرمی با قلبهای چهار حفرهای هستند. با این حال، آنها بیشتر با خزندگان مرتبط هستند و اعتقاد بر این است که از دایناسورها تکامل یافته اند. بیشتر پرندگان شبها میخوابند. بیشتر آنها میتوانند پرواز کنند، اما گونههای بدون پرواز وجود دارند.

بی مهرگان
در حالی که مهرهداران فقط از یک شاخه تشکیل شدهاند، بیمهرگان شاخههای باقیمانده از فرمانرو جانوران را تشکیل میدهند. همانطور که از نام آن پیداست، بیمهرگان فاقد ستون فقرات و اسکلت داخلی هستند. برخی از گونهها دارای یک سیستم اسکلتی خارجی به نام اسکلت بیرونی هستند که پشتیبانی ساختاری را فراهم میکند. در حال حاضر، بیمهرگان بیش از 97 درصد از تمام حیوانات در قلمرو حیوانات را تشکیل میدهند. بیمهرگان با داشتن چنین گروه متنوعی بر اساس تعدادی ویژگی از جمله مورفولوژی/ساختار، تقارن و چرخه زندگی و غیره طبقهبندی میشوند.
در حالی که برخی از موجودات دارای سه لایه بدن هستند، برخی دیگر (بیمهرگان اولیه) فقط دو لایه بدن دارند. نمونههایی از حیوانات بیمهره شامل حشرات (مانند عنکبوت و کنه)، حلزونها، ستارههای دریایی، اسفنجها، و هیدرها و غیره هستند.
گیاهان
این گروه از موجودات یوکاریوتهای چندسلولی هستند. سلولهای آنها دارای دیوارههای سلولی، واکوئلهای بزرگ اما فاقد سانتروزوم هستند. دیواره سلولی معمولاً از سلولز، پکتین و همیسلولز تشکیل شده است. آنها شیوه تغذیه متنوعی را نشان میدهند. اکثر آنها دارای کلروفیل a و b و رنگدانههای دیگر هستند، بنابراین معمولاً فتوسنتزی یا اتوتروف هستند. برخی مانند گیاهان حشرهخوار هتروتروف هستند.

گیاهان غیرمتحرک هستند، به این معنی که معمولاً در یک مکان ثابت میشوند. آنها دارای کلروپلاست هستند که اندامکی است که از جلبکهای تکسلولی تکامل یافته است. آنها به نمکهای معدنی، نور، اکسیژن و دیاکسیدکربن نیاز دارند. گیاهان این منابع را از محیط زیست به دست میآورند. نور و دیاکسیدکربن برای انجام فرآیند فتوسنتز ضروری هستند. بیشتر مواد غذایی خود را به شکل نشاسته ذخیره میکنند. بیشتر آنها از طریق جنسی، غیرجنسی و رویشی تولید مثل میکنند. عواملی مانند باد، هوا و حشرات به این فرآیند کمک میکنند. برخی از اعضای فرمانرو گیاهان تغییر نسل، یعنی فاز اسپوروفیت و فاز گامتوفیت را نشان میدهند. اعضای فرمانرو گیاهان سیستم دفعی و عصبی ندارند.
انواع گیاهان
گیاهان را میتوان با استفاده از معیار اولیه گلدهی به دوگروه گیاهان گلدار و بدون گل تقسیم کرد. گیاهان گلدار گل تولید میکنند و دانههای آنها در یک تخمدان قرار میگیرند. به گیاهان گلدار آنژیوسپرم (نهاندانگان) گفته میشود. آنژیوسپرمها بیشتر به تکلپهای و دولپهای تقسیم میشوند. جنین تکلپهای یک لپه دارد در حالی که جنین دولپهای دارای دو لپه است. نمونههایی از تک لپهها عبارتند از: ذرت، علف، گندم و نیشکر. نمونههایی از دولپهها عبارتند از لوبیا، گوجهفرنگی و چای.
گیاهان غیرگلدار گل تولید نمیکنند. در عوض، آنها میتوانند هاگ را در ساختارهای تخصصی یا دانههای برهنه روی سطح برگ خود تولید کنند. گیاهان غیرگلدار که دانههای برهنه تولید میکنند به عنوان گیاهان ژیمنوسپرم (بازدانگان) یا مخروطدار نامیده میشوند. بیشتر آنها درختان و درختچهها هستند و از نمونههای آن میتوان به کاج، صنوبر و سرو اشاره کرد. گیاهان غیرگلدار که اسپور تولید میکنند شامل اعضای تقسیم بندی «خزهها» (Bryophyta) و «سرخسها» (Pteridophyta) میشوند. بدن گیاهی خزهها به ساقه، ریشه و برگ تمایز نمییابد. در عوض، آنها فاقد ریشه بوده، اما دارای ساختارهای تخصصی به نام ریزوئید هستند که برای لنگر انداختن و جذب آب استفاده میشود. نمونههایی از بریوفیتها عبارتند از خزه و جگرواش.
از طرف دیگر، سرخسها دارای ریشه و سیستم ساقه هستند. علاوه بر این، آنها دارای یک سیستم آوندی با برگهای مرکب هستند که به نام برگ شناخته میشود. نمونههایی از پتریدوفیتها عبارتند از سرخسها و دماسبیان. هم خزهها و هم سرخسها تغییرات نسلی را نشان میدهند. اسپوروفیت گیاه اصلی در سرخسها است. در خزهها اسپوروفیت معمولاً به گامتوفیت متصل میشود.
قارچ ها
قارچ گروهی از ارگانیسمهای یوکاریوتی، غیرفتوتروف با دیوارههای سلولی سفت و سخت است که شامل قارچ، کپک و مخمر میشود. غیر فوتوتروف به این معنی است که آنها نمیتوانند از نور برای انرژی استفاده کنند زیرا کلروفیل ندارند و همین ویژگی آنها را از گیاهان متمایز میکند. بیشتر قارچها چندسلولی بوده و دارای دیوارههای سلولی هستند. آنها بیتحرک بوده، به این معنی که معمولا در یک مکان ثابت میمانند.

انواع قارچ ها
دانشمندان بیش از 100000 گونه قارچ را توصیف کردهاند. گونههای قارچی که شرح داده شده اند به پنج گروه طبقهبندی میشوند. نامها دشوار هستند، اما ممکن است متوجه شوید که هر گروه به پسوند «میست» (Mycetes) ختم میشود. بنابراین، اگر این پسوند را ببینید، متوجه میشوید که جاندار پیش رو قارچ است. در ادامه ۵ گروه مختلف قارچی را توضیح دادهایم:
- اولین گروه «قارچهای انگلی» (Chytridiomycetes) هستند که تصور میشود قدیمیترین گروه قارچ هستند. ما اغلب این قارچها را در دریاچهها، برکهها، مدخل رودخانهها و خاک پیدا میکنیم. بسیاری از گونههای این گروه تجزیهکننده هستند، به این معنی که مواد آلی مرده را تجزیه میکنند. اما گونههای دیگر انگلهای گیاهی و جانوری هستند و مسئول بسیاری از عفونتها در دوزیستان هستند.
- دومین گروه «زیگومیستها» (Zygomycetes) هستند که احتمالاً در مقطعی با آنها تعامل داشتهاید. برخی از قارچهای این گروه، کپکهایی هستند که به سرعت رشد میکنند، مانند کپکهای نان سیاه و کپکهایی که میوهای را که به مدت طولانی بیرون گذاشتهاید، میپوسند.
- «گلومرومیستها» (Glomeromycetes) که قارچهای بسیار مهمی هستند. این به این دلیل است که حدود 90 درصد گیاهان با گلومرومیستها روابط همزیستی برقرار میکنند که به رساندن مواد مغذی به گیاه کمک کرده و در عوض سایر مواد مغذی لازم را از آنها دریافت میکنند. مانند «کنگلومرا» که چندین بخش با هم ترکیب شده است، گلومرومیستها با گیاهان ترکیب میشوند تا یک تیم قوی را تشکیل دهند.
- آسکومیستها همچنین به عنوان قارچ «کیسهای» شناخته میشود. این قارچها نام خود را از ساختار کیسهمانند خود به نام «آسک» گرفته اند. اینها در زیستگاههای متنوعی (دریایی، آب شیرین و خشکی) زندگی میکنند و میتوانند از نظر اندازه از مخمر تکسلولی گرفته تا قارچهای مورل فانتزی متفاوت باشند. برخی از آسکومیستها میتوانند از مخربترین پاتوژنهای گیاهی باشند.
- «بازیدیومیستها» (Basidiomycetes)، همچنین به عنوان قارچ کانونی شناخته میشود. این همان چیزی است که اکثر مردم با شنیدن کلمه «قارچ» تصور میکنند زیرا بسیاری از آنها حالت چتری داشته و در زیر خود دایرههای متحدالمرکز دارند. بسیاری از این قارچها تجزیهکننده هستند، مانند قارچهای قفسهای که چوب را در جنگلها تجزیه میکنند. اما آنها همچنین شامل بسیاری از انگلهای گیاهی ویرانگر نیز میشوند.









