



تولید مثل باکتری ها چگونه است؟ – به زبان ساده
روش اصلی تولید مثل باکتری تقسیم دوتایی است که در این فرآیند سلول منفرد یک باکتری، به دو سلول دختری یکسان تقسیم میشود. آغاز تقسیم دوتایی با همانندسازی DNA است. روشهای متعدد دیگری نیز در باکتریهای مختلف برای تولید مثل وجود دارد که در این مطلب آنها را توضیح دادهایم.

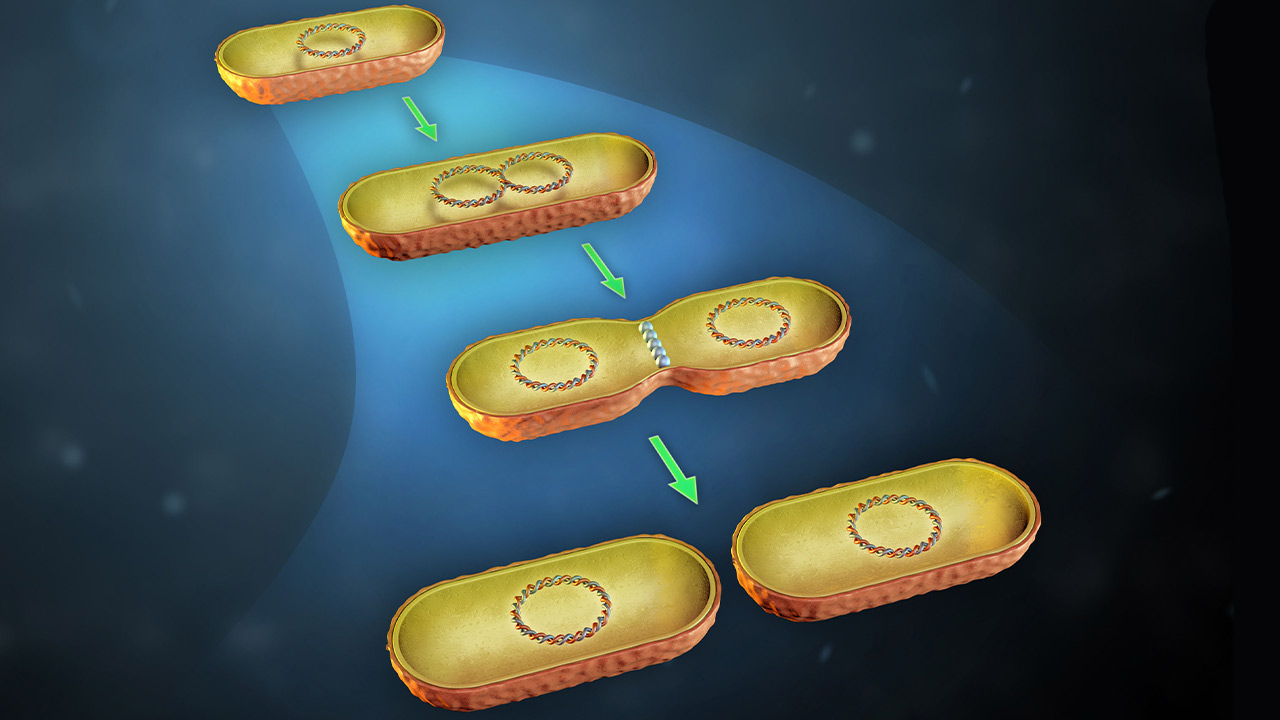


روش تولید مثل باکتری ها چیست؟
بیشتر باکتریها برای تکثیر به تقسیم دوتایی (Binary fission) متکی هستند که فرایندی ساده است. برای انجام تقسیم دوتایی باکتری باید ابتدا به اندازه دو برابر اندازه اولیه خود رشد کند. اما برای باقی ماندن و رقابت، یک باکتری باید در زمان مناسب و مکان مناسب تقسیم شود و یک نسخه کامل از مواد اساسی ژنتیکی خود را در اختیار سلولهای حاصل قرار دهد. تولید مثل باکتری در بسیاری از تحقیقات برای کشف مکانیسمهای ژنتیکی که تولید مثل باکتری را تنظیم و هدایت میکنند، مورد مطالعه قرار گرفتهاند.
درک چگونگی مراحل و مولکولی مؤثر در این فرآیند بسیار مورد توجه است چون امکان دارد از این طریق بتوان مواد شیمیایی جدید یا آنتیبیوتیکهای جدیدی تولید کرد که اختصاصا تولید مثل باکتری را هدف قرار داده و آن را مختل کنند. قبل از تقسیم دوتایی، سلول باید مواد ژنتیکی خود (DNA) را کپی کرده و این نسخهها را در انتهای مخالف سلول تفکیک کند. سپس انواع مختلفی از پروتئینها در محل تقسیم آینده جمع میشوند. جزء اصلی این سیستم، پروتئین FtsZ است. مونومرهای پروتئین FtsZ به صورت ساختاری شبیه حلقه در مرکز سلول جمع میشوند.
سپس سایر اجزای دستگاه تقسیم در حلقه FtsZ مونتاژ میشوند. این ماشین به گونهای قرار گرفته است که شکافت سیتوپلاسم، به DNA آسیب نمیرساند. با انجام تقسیم، سیتوپلاسم به دو بخش تقسیم و در بسیاری از باکتریها، همزمان دیواره سلولی جدیدی ساخته میشود. ترتیب و زمان فرآیندهای مختلف تولید مثل باکتری به روش دوتایی (همانندسازی DNA، تفکیک DNA، انتخاب محل تقسیم، معکوس شدن پاکت سلول و سنتز دیواره سلولی جدید) با مکانیسمهای متعددی کنترل میشوند.

مراحل تقسیم دوتایی باکتری
مانند سلول انسانی، تولید مثل باکتری وابسته به کپی کردن DNA است. بر خلاف سلولهای انسانی که دارای کروموزومهای محصور در یک هسته دارای غشا هستند، سلولهای باکتریایی معمولاً دارای یک کروموزوم حلقوی منفرد بوده و فاقد هسته هستند. با این حال، کروموزوم باکتری کاملا در سیتوپلاسم رها شده نیست بلکه در منطقه خاصی از سلول به نام نوکلوئید قرار دارد. همانندسازی DNA توسط آنزیمهای همانندسازی در نقطهای از کروموزوم به نام مبدأ همانندسازی آغاز میشود و در دو جهت مخالف ادامه مییابد. با ادامه تکثیر، دو منشأ به سمت انتهای مخالف سلول حرکت میکنند و کروموزوم را به همراه خود میکشند. سلول نیز بزرگتر میشود و به جدایی کروموزومهای تازه تشکیل شده کمک میکند.
همانندسازی تا زمانی که کل کروموزوم کپی شود ادامه مییابد و آنزیمهای همانندسازی در قسمت دورتر به هم میرسند. هنگامی که کروموزومهای جدید به انتهای سلول مخالف منتقل شدند، تقسیم سیتوپلاسم میتواند اتفاق بیفتد. در این فرآیند، غشا به داخل فشرده میشود و یک سپتوم (تیغه) یا دیواره جدید در وسط سلول تشکیل میشود. باکتریها دیواره سلولی دارند بنابراین وقتی تقسیم میشوند باید این دیواره را هم بازسازی کنند. سرانجام تیغه از وسط تقسیم شده و دو سلول آزاد میشوند تا به عنوان باکتریهای جداگانه به زندگی خود ادامه دهند.

تفاوت تقسیم دوتایی با میتوز چیست؟
تقسیم دوتایی باکتری از برخی جهات شبیه تقسیم میتوز در انسان و سایر یوکاریوتها است. در هر دو مورد، کروموزومها کپی و از هم جدا میشوند و سلول سیتوپلاسم خود را تقسیم کرده و دو سلول جدید تشکیل میدهند. با این حال، فرایند آنها نسبتاً متفاوت است. در تولید مثل باکتری هیچ دوکی از نوع میتوزی ایجاد نمیشود و مهمتر از همه، همانندسازی DNA همزمان با جداسازی DNA طی تقسیم دوتایی اتفاق میافتد، درحالی که در تقسیم میتوز، DNA در طول فاز S چرخه سلولی همانندسازی میشود که قبل از شروع تقسیم و ورود به مرحله میتوز است.
انواع روش تولید مثل باکتری چه هستند؟
تولید مثل باکتری انواع غیرمعمول یا الگوهای تقسیم سلولی دیگری نیز دارد که در برخی گونهها یا شرایط خاص اتفاق میافتند. برخی از باکتریها بیش از دو برابر اندازه سلول اولیه رشد میکنند و سپس با استفاده از تقسیمات متعدد، چندین سلول جدید تولید میکنند. برخی از دودمانهای باکتریایی با جوانه زدن تولید مثل میکنند. تولید مثل باکتری میتواند از طریق ایجاد یک سلول جدید در داخل سیتوپلاسم سلول والد باشد. در ادامه انواع رایجتر تولید مثل باکتری به روشهای غیر معمول را توضیح دادهایم.
هم یوغی چیست؟
هم یوغی باکتری (Bacterial Conjugation) یکی از سه حالت اصلی شناخته شده تبادل ژنتیکی بین باکتریها است و دو حالت دیگر انتقال و تبدیل باکتری هستند. از بین این سه حالت، تنها حالت است که شامل تماس سلول به سلول است. «J. Lederberg» و «E. L. Tatum» برای اولین بار چنین انتقالی را در سال 1946 در اشرشیا کولی (E. coli) گزارش دادند. این کشف در نتیجه جستجوی نوترکیبی جنسی در باکتریها حاصل شد که در آن سلولهای دختری، مارکرهای ژنتیکی را از دو والد حمل میکنند.
هم یوغی باکتری یک روش انتقال ژنتیکی جنسی است به این معنا که مواد کروموزومی از دو نوع سلول متمایز از نظر جنسی در یک فرآیند مشخص و برنامهریزیشده در سلول دختری گرد هم میآیند. با این حال، در مقایسه با تولید مثل جنسی در بیشتر ارگانیسمهای پیشرفته که شامل تعامل بین کل مجموعه کروموزومها از هر دو نوع سلول والدی است، در هم یوغی این فرآیند تنها بخشی از ژنوم اهدا کننده و ژنوم گیرنده را در بر میگیرد.
بنابراین در هم یوغی باکتریها انتقال ژنتیکی به صورت جزئی است و در اکثر موارد حالت قطبی دارد، یعنی مواد ژنتیکی به طور یک طرفه از سلول اهدا کننده به سلول گیرنده منتقل و به دنبال آن سلولهای جدید از یکدیگر جدا میشوند و تغییرات بیشتری در سازمان یا نوترکیبی مواد ژنتیکی درون سلول گیرنده ایجاد خواهد شد. انتقال مواد ژنتیکی ممکن است چند دقیقه یا بیشتر (تا چند ساعت) طول بکشد. بعدها مشخص شد که عامل این فرآیند، ناحیهای در کروموزوم است که فاکتور F (باروری) نامیده میشود.
این یافته اساس ژنتیک باکتریها در دهههای 1940 و 1950 بود و به طور گستردهای در نقشهبرداری از کروموزوم E. coli مورد استفاده قرار گرفت و آن را به ارگانیسم برجسته پروکاریوتی در آن زمان تبدیل کرد. همچنین نشان داده شد که فاکتور F میتواند از کروموزوم خارج شود و به عنوان یک عنصر یا پلاسمید خارج کروموزومی وجود داشته باشد. این ماده قادر به انتقال به سایر باکتریها بوده و میتواند به طور تصادفی در DNA میزبان خود ادغام شود. فاکتور جنسی F باکتری E. coli، حساسیت را به باکتریوفاژهایی که به پیلوس F یا «F pilus» نیاز دارند منتقل میکنند که به عنوان محل اتصال در هنگام عفونت، توسط ناحیه F رمزگذاری میشود.
در دهه 1960 تعداد دیگری از پلاسمیدهای پیوندی جداسازی شدند که بسیاری از آنها دارای مارکرهای مقاومت آنتیبیوتیکی متعددی بودند. این پلاسمیدها فاکتورهای R (مقاومت) نامیده میشوند و در بسیاری از موارد برای سرکوب همانندسازی و ترکیب با فاکتور F (فرآیندی که مهار باروری نامیده میشود) یافت شدند. تعداد پلاسمیدهای پیوندی کشف شده در چند دهه گذشته بسیار رشد کرده است و شامل پلاسمیدهایی است که به صورت مستقل قابل انتقال هستند و از باکتریهای گرم منفی، گرم مثبت و همچنین پلاسمیدهای متحرک جدا شدهاند.

ترانسپوزونهای همجوشی یا عناصر همجوشی یکپارچه (ICE) که با استفاده از مکانیسم پیوندی بین سلولها حرکت میکنند، از طریق فرآیندی که باعث یادآوری فاژهای لیزوژنیک میشود، از بدن جدا شده و در کروموزوم میزبان قرار میگیرند. یک نمونه از فاژهای پیوندی برای استافیلوکوکوس اورئوس توصیف شده است. به طور کلی، توابع انتقال و تکثیر این عناصر متحرک، غالباً از نظر فیزیکی به هم پیوند میخورند. هم یوغی باکتری یکی از اصلیترین کانالهای انتقال افقی ژن (HGT) در میان میکروارگانیسمها است.
این فرآیند بسیار گسترده بوده و میتواند از طریق ژن و همچنین بین فرمانروها (باکتری به مخمر یا گیاه) رخ دهد. تجزیه و تحلیل توالی DNA باکتریها نشان داده است که اتصال و تغییر (دو کانال اصلی برای انتقال افقی ژن)، توسط یک مجموعه پروتئین متعلق به سیستم ترشحی نوع IV انجام میشوند. این فرآیند تأثیر زیادی بر تکامل بسیار داشته، زیرا باکتریها به سرعت صفات خوب (استفاده از هیدروکربن) و بد (مقاومت آنتیبیوتیکی، سموم) را به دست میآورند. تولید مثل باکتری با هم یوغی یکی از مباحث مهم میکروبیولوژی است (با تأکید بر خود فرآیند نه صرفا کاربرد آن به عنوان یک ابزار ژنتیکی).
میتوان پیشنهاد کرد که دو سویه باکتری واقعاً ژن رد و بدل نمیکنند بلکه در عوض مواد نشتی دارند و سلولهای دیگر میتوانند آن را جذب کرده و از آنها برای رشد استفاده کنند. احتمال تغذیه متقابل توسط برنارد دیویس رد شد. او یک لوله U ساخت که در آن دو بازو با یک فیلتر خوب از هم جدا شدند. منافذ فیلتر برای عبور باکتری بسیار کوچک بودند اما به اندازه کافی بزرگ بودند که امکان عبور راحت محیط مایع و هرگونه مواد محلول را فراهم میکردند. سویه A در یک بازو قرار گرفت و سویه B در دیگری.
بعد از مدتی انکوباسیون سویهها، دیویس محتوای هر بازو را آزمایش کرد تا ببیند سلولها قادر به رشد در حداقل محیط هستند یا خیر. به عبارت دیگر، تماس فیزیکی بین دو سویه برای تشکیل سلولهای نوع وحشی مورد نیاز بود. به نظر میرسید که نوعی انتقال ژن اتفاق افتاده است و در واقع نوترکیبهای ژنتیکی تولید میشوند. انتقال ژنتیکی در یک جهت در انواع مختلف تلاقی رخ داده است. بنابراین، انتقال مواد ژنتیکی در E. coli متقابل نیست. یک سلول به عنوان اهدا کننده و سلول دیگر به عنوان گیرنده عمل میکند. این نوع انتقال یک طرفه ژنها در اصل با تفاوت جنسی مقایسه میشد، در حالی که اهدا کننده پدری و گیرنده مادری نامیده میشد.

با این حال، این نوع انتقال ژن تولید مثل جنسی واقعی نیست. در انتقال ژن باکتریایی، یک ارگانیسم اطلاعات ژنتیکی را از اهدا کننده دریافت میکند و گیرنده توسط آن اطلاعات تغییر مییابد. در تولید مثل جنسی باکتری، ماده ژنتیکی هر دو ارگانیسم به طور مساوی (یا تقریباً مساوی) به ارگانیسم جدید اهدا میشوند اما فقط در موارد استثنایی هریک از اهداکنندگان تغییر میکند. باروری (توانایی اهدا) E. coli به راحتی از دست میرود و دوباره به دست میآید. توانایی اهدا کنندگی یک حالت وراثتی است که توسط عامل باروری (F) حمل میشود. سویههای حامل فاکتور F میتوانند اهدا کننده باشند بنابراین +F گفته میشوند. سویههایی که فاقد فاکتور F هستند نمیتوانند اهداکننده باشند بلکه گیرنده هستند و −F نامیده میشوند.
ژنوتیپهای نوترکیب برای ژنهای مارکر در تلاقیهای باکتریایی نسبتاً نادر هستند اما فاکتور F ظاهراً در اثر تماس یا جفت شدن منتقل میشود. به نظر میرسد نوعی انتقال عفونی فاکتور F در حال انجام است. اکنون اطلاعات بیشتری در مورد روند ترکیب و F وجود دارد که نمونهای از یک پلاسمید است که میتواند به طور مستقل از کروموزوم میزبان در سیتوپلاسم تکثیر شود. پلاسمید F سنتز پیلی را هدایت میکند، تماس با یک گیرنده، به DNA F اجازه میدهد از طریق منافذ وارد سلول گیرنده شود.
یک رشته از DNA دو رشته F منتقل میشود و سپس همانندسازی DNA رشته مکمل را در اهدا کننده و گیرنده بازیابی میکند. این همانند سازی باعث میشود که کپی F در اهدا کننده باقی بماند و دیگری در گیرنده ظاهر شود. برخی گونههای مشتق شده از سویه +F در هم یوغی با سویه −F که 1000 برابر تعداد بیشتری نوترکیب برای مارکرهای ژنتیکی تولید کرده است که سویه Hfr نام دارد. در عبورهای Hfr و −F، تقریباً هیچیک از والدین −F به +F یا Hfr تبدیل نمیشوند. در هم یوغی بین +F و −F انتقال عفونی F منجر به تغییر بخش زیادی از والدین −F به +F میشود.
یک سویه Hfr از ادغام فاکتور F در کروموزوم حاصل میشود. در طی اتصال بین سلول Hfr و سلول −F، بخشی از کروموزوم با F. منتقل میشود شکستگی تصادفی انتقال را قبل از انتقال کل کروموزوم قطع میکند. سپس قطعه کروموزومی میتواند با کروموزوم گیرنده ترکیب شود. واضح است که سطح پایین انتقال نشانگر کروموزومی در هم یوغی بین سویههای +F و −F را میتوان با وجود سلولهای نادر Hfr در جمعیت توضیح داد. هنگامی که این سلولها جدا و خالص میشوند، مارکرهای کروموزومی را با فرکانس بالا منتقل میکنند، زیرا هر سلول یک Hfr است.

توانایی باکتریهای بیماریزا در بیمارستانهای ژاپن در دهه 1950 کشف شد. اسهال خونی باکتریایی توسط باکتریهای جنس شیگلا ایجاد میشود. این باکتری در ابتدا نسبت به مجموعه وسیعی از آنتیبیوتیکهایی که برای کنترل بیماری استفاده میشدند، حساس بود. با این حال، در بیمارستانهای ژاپن ثابت شد شیگلا جدا شده از بیماران مبتلا به اسهال خونی در برابر بسیاری از این داروها از جمله پنیسیلین، تتراسایکلین، سولفانیل آمید، استرپتومایسین و کلرامفنیکل به طور همزمان مقاومت دارد.
فنوتیپ مقاوم در برابر داروهای متعدد، به عنوان یک بسته ژنتیکی منفرد به ارث رسیده است و میتواند به روش عفونی نه تنها به سایر سویههای حساس شیگلا، بلکه به سایر گونههای مرتبط باکتری نیز منتقل شود. این استعداد برای باکتری بیماری زا فوقالعاده مفید و پیامدهای آن برای علوم پزشکی وحشتناک است. پلاسمید حامل این مقاومتها یک عنصر خودتکرار شبیه به فاکتور F است. فاکتورهای R دقیقاً مانند ذره F در E. coli، به سرعت در هم یوغی منتقل میشوند. در حقیقت، این فاکتورهای R تنها اولین فاکتور مشابه F مانند کشف شده هستند. این عناصر که به صورت پلاسمید در سیتوپلاسم وجود دارند، انواع مختلفی از ژنها را در باکتریها حمل میکنند.

کروموزوم Hfr در حالی که یک رشته را به سلول −F منتقل میکند، همانندسازی میشود و وجود یک کروموزوم کامل را برای سلول اهدا کننده پس از هم یوغی و انتقال تضمین میکند. رشته منتقل شده در سلول گیرنده تکثیر میشود و ژنهای دهنده از طریق کراسینگ اوور در کروموزوم گیرنده قرار میگیرند و سلول نوترکیبی ایجاد میکنند. در غیر این صورت، قطعات منتقل شده DNA در گیرنده در طی تقسیم سلول از بین میروند.
فرض میکنیم که کروموزوم −F نیز حلقوی است، زیرا اگر سلول گیرنده فاکتور F را از سلول +F دریافت کند، به راحتی به سلول +F تبدیل میشود که میتوان از آن سلول Hfr استخراج کرد. سلولهای Hfr و +F دارای ساختارهای فیبری به نام پیلی F هستند که از دیواره سلولی آنها بیرون زده است. F پیلی تماس سلول با سلول را تسهیل میکند که طی آن DNA از طریق منافذ در −F منتقل میشود.
تولید مثل باکتری با کیست
بعضی از باکتریها مانند سیانوباکتریوم استانیریا از طریق تقسیم دوتایی تولید مثل نمیکنند، بلکه تولید مثل باکتری از یک سلول کوچک و کروی شروع میشود که قطر آن تقریباً 1 تا 2 میکرومتر است. از این سلول به عنوان بائوسیت (که در لغت به معنای سلول کوچک است) یاد میشود. بائوسیت شروع به رشد میکند و در نهایت یک سلول رویشی با قطر 30 میکرومتر به وجود میآید. با رشد، DNA سلولی بارها همانندسازی میشود و سلول یک ماتریکس خارجی ضخیم تولید میکند.
سلول رویشی در نهایت به یک مرحله تولیدمثلی تبدیل میشود که در آن تعدادی شکاف سیتوپلاسمی وجود دارند تا به دهها یا حتی صدها سلول تقسیم شوند. ماتریس خارج سلولی سرانجام پاره میشود و سلولهای جدید را آزاد میکند. سایر اعضای راسته Pleurocapsales (یک راسته از سیانوباکتریوم) نیز از الگوهای غیر معمول تقسیم استفاده میکنند.

تولید مثل باکتری با جوانه زدن
جوانه زدن (Budding) در برخی از اعضای باکتریهای پلانکتومیستها، سیانوباکترها، فیرمیکوتها (باکتریهای گرم مثبت با محتوای G + C اندک) و پروتئوباکتریا مشاهده میشود. اگرچه جوانه زدن به طور گسترده در مخمر یوکاریوتی ساکارومیسس سرویزیه مورد مطالعه قرار گرفته است اما مکانیسمهای مولکولی تشکیل جوانه در باکتریها مشخص نیست.
باکتریهای جوانه زده با قرار گرفتن مواد جدید در یک نقطه از دیواره سلول رشد میکنند. شواهد فزایندهای وجود دارد که نشان میدهد این فرایند در بین انواع باکتریها رایج است و پیامدهای این نوع رشد از اهمیت زیادی برخوردار است. قطبیت سلول از ویژگیهای روند جوانه زدن باکتری است. هنگام تقسیم دو سلول غیر یکسان تولید میشوند که یکی از آنها زنده مانده و تکثیر میشود.

تولید مثل درون سلولی باکتری
تولید مثل مایکوباکتریوم پلیاسپورا (Metabacterium polyspora)، اپیولوپیسوم فیشلسونی (Epulopiscium spp) و باکتریهای رشتهای قطعه قطعه (SFB) سلولهای جدید متعددی را درون درون سلول تولید میکند. به نظر میرسد برای برخی از این باکتریها، این روند تنها راه تولید مثل است. رشد درون سلولی در این باکتریها ویژگیهای مشترکی با تشکیل اندوسپور در باسیلوس سوبتیلیس دارد. در گونههای اپیولوپیسوم این استراتژی تولید مثل باکتری با تقسیم نامتقارن سلول آغاز میشود.
به جای قرار دادن حلقه FtsZ در مرکز سلول، همانند تقسیم دوتایی، حلقههای Z در نزدیکی هر دو قطب سلول در اپیولوپیسوم قرار میگیرند. تقسیم سلول اولیه بزرگ، دو سلول دختری کوچک تشکیل میدهد که حاوی DNA هستند و به طور کامل توسط سلول والدی احاطه شدهاند. سلولهای دختری درون سیتوپلاسم سلول مادر رشد میکنند. هنگامی که تکامل سلولهای دختری کامل شد، سلول والد میمیرد و سلولهای دختری آزاد میشوند.
تولید مثل باکتری با کنیدیا
تشکیل کنیدیا (Conidia) در باکتریهای رشتهای مانند استرپتومایسس، با تشکیل یک تیغه عرضی در رأس رشته رخ میدهد. بخشی از این رشته را که دارای کویدیا است، کندیوفور مینامند. پس از جدا شدن از سلول مادری و تماس با بستر مناسب، كنیدیم جوانه زده و باعث ایجاد میسلیوم جدید میشود.

تولید مثل باکتری با اسپور
اسپورها در شرایط نامساعد محیطی مانند خشک شدن و نبود مواد مغذی تشکیل میشوند. از آنجا که اسپور در داخل سلول تشکیل میشود، آنها را آندوسپور مینامند. فقط یک اسپور در سلول باکتریایی و در هنگام جوانه زدن، یک سلول باکتریایی ایجاد میشود. برخی از باکتریهای تشکیلدهنده اندوسپور عبارتند از:
- باسیلهای گرم مثبت:
- باکتریهای هوازی مثل Bacillus subtilis و B. anthracis
- باکتریهای بیهوازی مثل کلستریدیوم تتانی و بوتولینوم
- کوکسیهای گرم مثبت مثل Sporosarcina
- باسیلوسهای گرم منفی مثل Coxiella burnetii
- کوکسیهای گرم منفی مانند اشرشیا کولی

روند تشکیل آندوسپور پیچیده است. ارگانیسم مدل مورد استفاده برای مطالعه تشکیل اندوسپور، باسیلوس سوبتیلیس است. تکمیل اندوسپور به چندین ساعت زمان نیاز دارد. تغییرات مورفولوژیکی کلیدی در فرآیند به عنوان مارکر برای تعیین مراحل رشد استفاده میشوند. با آغاز فرایند، سلول به صورت نامتقارن تقسیم (مرحله II) و دو محفظه شامل سلول مادر بزرگتر و سلول کوچکی ایجاد میکند. این دو سلول سرنوشت رشدی متفاوتی دارند. سیستمهای ارتباطی بین سلولی بیان ژن، سلول خاص را از طریق فعالسازی پی در پی فاکتورهای خاص سیگما در هریک از سلولها هماهنگ میکنند.
در مرحله بعد (مرحله III)، پپتیدو گلیکان در تیغه تخریب و پیش ماده توسط سلول مادر غرق میشود و سلول دختری را در داخل سلول تشکیل میدهد. فعالیتهای سلول مادری و پیش ماده منجر به سنتز ترکیبات خاص آندوسپور، تشکیل قشر و رسوب پوشش میشوند (مراحل IV + V). به دنبال آن کم آبی و بلوغ نهایی آندوسپور (مراحل VI + VII) اتفاق میافتند. سرانجام سلول مادری در اثر مرگ برنامهریزی شده سلول تخریب و آندوسپور در محیط آزاد میشود. اندوسپور تا زمانی که بازگشت شرایط مساعدتر را حس نکند، خاموش خواهد ماند. (فاکتور سیگما یک پروتئین کوچک است که RNA پلیمراز را برای شروع بیان ژن به مناطق خاص موجود در DNA هدایت میکند).
برخی از باکتریهای همزیست با ماهی مانند اپیولوپیسوم، آندوسپور بالغ تشکیل میدهند. این اسپورها دارای تمام لایههای محافظی هستند که در اندوسپورهای باسیلوس سابتیلوس (B. subtilis) دیده میشوند و همچنین حاوی مقادیر زیادی اسید دیپیکولینیک هستند. اینها با اندازهای بیش از 4000 برابر اندوسپور باسیلوس سوبتیلیس، بزرگترین اندوسپورهایی هستند که تاکنون توصیف شدهاند. تشکیل اندوسپورها ممکن است به حفظ ارتباط همزیستی بین این همزیستهای قارچ مانند و میزبانان جراح ماهی آنها کمک کند.
از آنجا که تشکیل آندوسپور همزمان با دوره هایی است که جراح ماهی میزبان به طور فعال تغذیه نمیکند، سلولها نیازی به رقابت برای مواد مغذی محدود موجود در روده در شب ندارند. خواص محافظتی آندوسپورها همچنین به آنها امکان میدهد تا از میزبانهای جدید جراح ماهی زنده بمانند. ماهی همچنین ممکن است از این رابطه بهرهمند شود زیرا قادر به حفظ جمعیت پایدار میکروبی است که به هضم غذا کمک میکند و ممکن است از محصولات میکروبی آزاد شده در هنگام مرگ سلولهای مادر و جوانه زدن اسپورها، سود غذایی دریافت کند.

مراحل رشد باکتری چه هستند؟
تولید مثل باکتری در شرایط مساعد سرعت نمایی دارد و در محیط کشت، الگوی رشد کلونی قابل پیشبینی است. این الگو را میتوان به صورت گرافیکی به عنوان تعداد سلولهای زنده یک جمعیت در طول زمان نشان داد و به عنوان یک منحنی رشد باکتری شناخته میشود. چرخه رشد باکتری در یک منحنی رشد شامل چهار مرحله تأخیر، نمایی، ثابت و مرگ است.

باکتریها برای رشد به شرایط خاصی نیاز دارند و این شرایط برای همه باکتریها یکسان نیست. عواملی مانند اکسیژن، pH، دما و نور در رشد میکروبی تأثیر دارند. عوامل اضافی شامل فشار اسمزی، فشار اتمسفر و در دسترس بودن رطوبت است. زمان تولید یک جمعیت باکتریایی یا مدت زمان دو برابر شدن جمعیت، در گونهها متفاوت است و به میزان نیازهای رشد بستگی دارد.
مراحل منحنی رشد باکتری
در طبیعت باکتریها شرایط کاملی را برای رشد تجربه نمیکنند. به همین دلیل، گونههای غالب که در یک محیط معمولا با گذشت زمان تغییر میکنند. اما در آزمایشگاه با رشد باکتریها در محیط کشت بسته میتوان شرایط بهینه را تأمین کرد. تحت این شرایط است که میتوان الگوی منحنی رشد باکتری را مشاهده نمود. منحنی رشد باکتری نشاندهنده تعداد سلولهای زنده در یک جمعیت باکتری، طی یک دوره زمانی مشخص است.

- فاز تأخیر (Lag Phase): این فاز اولیه با فعالیت سلولی مشخص میشود اما رشد ندارد. گروه کوچکی از سلولها در محیط غنی از مواد مغذی قرار میگیرند که به آنها امکان میدهد پروتئینها و سایر مولکولهای لازم برای تکثیر را سنتز کنند. اندازه این سلولها افزایش مییابد اما هیچ تقسیم سلولی در مرحله رخ نمیدهد.
- فاز نمایی (Exponential (Log) Phase): پس از مرحله تأخیر، سلولهای باکتریایی وارد مرحله نمایی یا ورود به سیستم میشوند. این زمانی است که سلولها پس از هر بار تولید مثل باکتری ها با تقسیم دوتایی و دو برابر شدن تعداد تقسیم میشوند. فعالیت متابولیک به دلیل تولید DNA، RNA، اجزای دیواره سلولی و سایر مواد لازم برای رشد برای تقسیم، زیاد است. آنتیبیوتیکها و ضد عفونیکنندهها بیشترین تأثیر را در این مرحله رشدی دارند زیرا این مواد معمولاً دیواره سلولهای باکتری یا فرآیندهای سنتز پروتئین، رونویسی از DNA و ترجمه RNA را هدف قرار میدهند.
- فاز ثابت (Stationary Phase): با کاهش مواد مغذی موجود و شروع به تجمع مواد زائد، رشد جمعیتی که در مرحله ورود به سیستم تجربه میشود، شروع به کاهش میکند. رشد سلولهای باکتریایی به یک فلات یا فاز ساکن میرسد، جایی که تعداد سلولهای تقسیمکننده با تعداد سلولهای در حال مرگ برابر است. این منجر به عدم رشد کلی جمعیت میشود. در شرایط کمتر مطلوب، رقابت برای مواد مغذی افزایش مییابد و سلولها از نظر متابولیکی فعالیت کمتری دارند. باکتریهای تشکیلدهنده اسپور در این مرحله اندوسپورها را تولید میکنند و باکتریهای بیماریزا شروع به تولید موادی (عوامل حدت) میکنند که به آنها کمک میکند تا از شرایط سخت زنده بمانند و در نتیجه باعث بیماری شوند.
- فاز مرگ (Death Phase): با كمتر شدن مواد مغذی و افزایش مواد زائد، تعداد سلولهای در حال مرگ در حال افزایش است. در مرحله مرگ، تعداد سلولهای زنده به طور تصاعدی کاهش مییابد و رشد جمعیت کاهش چشمگیری را تجربه میکند. سلولهای در حال مرگ در حال لیز یا باز شدن، محتوای آنها به محیط ریخته میشود و این مواد مغذی را در دسترس سایر باکتریها قرار میدهد. این به باکتریهای تولیدکننده اسپور کمک میکند تا برای تولید اسپور به اندازه کافی زنده بمانند. اسپورها میتوانند در شرایط سخت مرحله مرگ زنده بمانند و با قرار گرفتن در محیطی که زندگی را پشتیبانی میکند، به باکتری در حال رشد تبدیل میشوند.










من در مورد فرمول نرخ رشد ویژه باکتریها سوال داشتم.