



هتروکرونی یا ناهمزمانی تکوینی–تکاملی – از صفر تا صد
تا به حال فکر کردهاید که بالهای خفاش چگونه از دستهای کوتاه و معمولی پستانداری موشمانند و غیرپروازگر تکامل یافته است؟ مثلا جهشی ناگهانی رخ داده و پستاندار موشمانند و غیرپروازگر ما، ناگهان بچههایی با انگشتان دراز و پردهای پوستی میان انگشتان زاییده است؟ یا مثلا چطور گردن زرافه تکامل یافته و از نسل جانوری با گردن متعارف (مثلا شبیه اوکاپی/Okapi یا همان زرافه گردنکوتاه) موجودی با گردن نامتعارف و بلند پیدا شده است؟




جواب خیلی کوتاه برای این پرسشها وجود ندارد؛ ولی اگر بخواهیم خیلی ساده ماجرا را تعریف کنیم به این صورت بوده که تکامل این صفات به صورت تدریجی رخ داده است. منتها چه فرایندی پشت این تکامل تدریجی بوده؟ حتما برایتان جالب خواهد بود که بدانید اتفاقا خیلی از صفات کوچک و بزرگ در جانوران مختلف، از شکل ملوس و دوستداشتنی گربهها بگیرید تا صورت بدون پوزهای که وجه تمایز انسان با سایر میمونهاست، همگی وابسته به همین فرایند تکاملی بوده. فرایندی که آن را هتروکرونی (Heterochrony) مینامیم و فهمیدن سازوکار آن مسائل زیادی را درباره چگونگی جانوران و گیاهان با ریختشناسیهای متنوع و گوناگون برای ما آشکار خواهد کرد.
تعریف و پیشینه
این اصطلاح مفهومی روشن و صریح دارد. این واژهی لاتین، برگرفته از دو ریشهی یونانی هتروس (Heteros / ἕτερος به معنی متفاوت / دیگر) و خرونوس (Khronos / χρόνος به معنی زمان) معنایی چون دیگرشدن زمانبندی میدهد. بهعبارت دیگر تعریف هتروکرونی، نهفته در ریشهی خود واژه است.
برای نخستینبار، ارنست هکل (Ernst Heinrich Philipp August Haeckel (February 16, 1834 – August 9, 1919)) این واژه را در سال 1875 بهکاربرد. از آن زمان تا کنون، هتروکرونی، بهعنوان یکی از مفاهیم اصلی زیستشناسی تکاملی درآمده است.

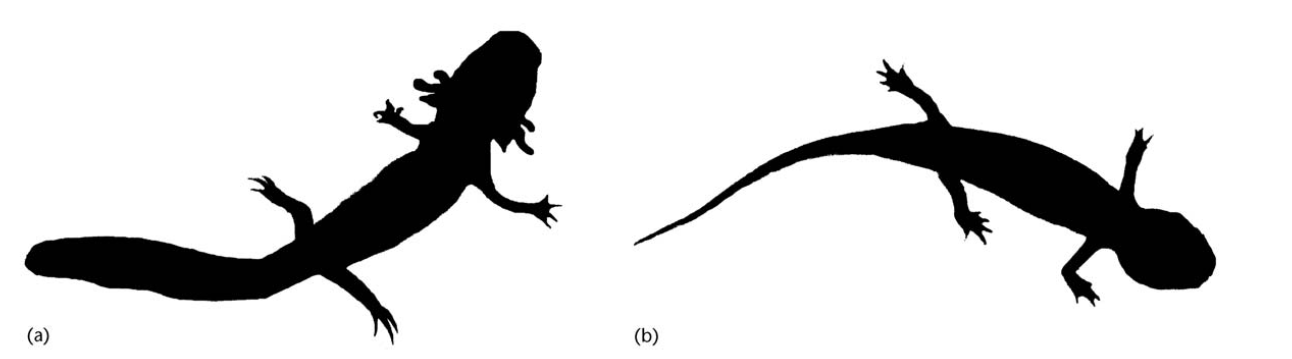
مفهوم دقیقتر هتروکرونی، تغییر در نرخ تکوین جنینی، در طی تکامل است. مثال بارز و آشنای آن نیز اکسیلاتل مکزیکی (Mexican Axolotl (Ambystoma mexicanum)) است که ظاهراً در اثر کندشدن و ابترشدن تکوین این جانور در طی تکامل، بالغ این گونهی امروزی، شکلی همانند لاروهای نیاکان خود پذیرفته است.
اکسیلاتل شناختهشدهترین گونه از سمندرهای کور و نئوتنیک مکزیکی، متعلق به گروه سمندرهای ببری است. لاروهای این گونه، قادر به دگردیسی نیستند و در زمان بلوغ، همچنان آبزی و آبششدار باقیمیمانند. این گونه بومی دریاچهای زیرزمینی در زیر مکزیکوسیتیست و در زبان مردم محلی آهولوته (Ajolote) بهمعنی موشکور نامیده میشود.
اهمیت هتروکرونی در تکامل فوقالعاده زیاد است. میتوانگفت بیشتر جنبههای تکامل انسان از طریق هتروکرونی قابل توجیههستند: بدن بزرگ، مغز بزرگ، طولانیشدن دورهی یادگیری و طول زندگی (Horder, 2006). با تکیه بر مفهوم تکرار رئوس (Recapitulation) هکل، میتوان گفت تغییر در نرخ تکوین جنینی، بر اثر تغییراتی تکاملی ظاهرمیشود. برای نخستینبار، هکل بود که ابرازکرد، سیر تکوین فردی، تکرار سریع تکامل نوعیست.
این مفهوم گرچه عمومیت ندارد، اما کمابیش درستاست. بنابراین در دههی نود قرن گذشته، هتروکرونی بهعنوان توضیح عمومی بسیاری از تغییرات تکاملی مطرحگردید (McNamara, 1990) (McNamara, 2003). با وجود این سادگی آغازین، زمانی که به شرایط ایجادکنندهی هتروکرونی و اقسام برآیندهای نهایی این پدیده بنگریم، دیگر آن را ساده نخواهیم یافت.
بنابراین میکوشیم «اقسام» هتروکرونی را طبقهبندیکنیم و ارتباطی تنگاتنگ میان این مفهوم و پدیدههایی چون نئوتنی (Neoteny)، پدومورفوز (Paedomorphosis) و از کار افتادگی (Vestigialization) بیابیم. مثال اکسیلاتل، مثالی خوب برای نئوتنیست. بهطور کلی نئوتنی برآمدی از هتروکرونیست که در نتیجهی آن افراد گونهی جدید به شکلی تغییرکردهاند که وقتی به بلوغ جنسی برسند، ظاهری چون افراد نابالغ گونههای نیایی مییابند.
این ظاهر نابالغ در اثر کندشدن تکوین و رشد اندامها نسبت به دستگاه تناسلیست. در مورد نئوتنی گفتهشد افراد بالغ گونهی دختری، ظاهری چون افراد نابالغ گونهی نیایی دارند و این بر اثر کندشدن نرخ رشد برخی اندامهاست. جز این ممکناست بدون تغییر در نرخ رشد، زمان توقف رشد برخی اندامها پیشتر بیافتد و افراد گونهی مزبور بازهم ظاهری نابالغ خواهندیافت. به این پدیده پروژنز (Progenesis) گفتهمیشود. هم پروژنز و هم نئوتنی، صورتی از پدومورفوز هستند.
پدومورفوز-فارغ از چگونگی بروز-کلاً به ظهور چهره و صفات کودکانه در افراد بالغ گونه گفتهمیشود. تحلیلرفتن و ازکارافتادگی برخیاندامها نیز طی تکامل در اثر تغییر نرخرشد یا پیشافتادن زمان توقف رشد آنها روی میدهد. بنابراین میتوان ازکارافتادگی اندامهایی چون رودهی کور (Appendix) یا پلک سوم را به هتروکرونی نسبتداد. به هرحال در طی زمانی که تا امروز از معرفی این فرایند میگذرد، تعاریف و مفاهیم گوناگونی برای هتروکرونی بیانشدهاند. بد نیست مروری به این تعاریف تاریخی داشته باشیم.
تعاریف تاریخی
در طی یکصد و بیست سال گذشته، هتروکرونی نقش مهمی در پیوند فیلوژنی (زیستشناسی تکاملی) و انتوژنی (زیستشناسی تکوینی) بازیکردهاست. بنابراین تعجبینخواهد داشت اگر در طی این مدت تعاریف بسیار متنوع و گستردهای میان زیستشناسان برای هتروکرونی درنظرگرفته شده باشد.
دورهی نخست (۱۸۶۶-۱۸۷۵)
در دوره نخست، تکوین جنین بهعنوان بازنمود دقیق وقایع تکاملی درنظرگرفتهمیشود. واژهی هتروکرونی پیش از هکل نیز وجود داشت (Horder, 2006) اما این هکل بود که مفهوم آن را تغییرداد و به الگوی امروزین نزدیکشد. او هترکرونی را به عنوان یکی از استثنائات قانون تکرار رئوس خود مطرحکرد، چراکه در اثر هتروکرونی، بخشی از دستاوردهای فیلوژنی در حین تکوین از میان میرود.
دورهی دوم (۱۸۷۵-۱۹۲۵)
دوره دوم، دوره انباشت مثالهای نقض برای قانون تکرار رئوس هکل. در این دوره که نقدهای شدیدی به قانون هکل واردمیشد، مفهوم هتروکرونی توسعهپیداکرد. این موضوع پذیرفتهشدهبود که تکوین هر اندام، تکرار تکامل همان اندام در گذشته است، فارغ از اینکه تکوین اندامها با نرخ تکوین کلی جنین هماهنگ باشد یا خیر.
بنابراین بر این موضوع صحهگذاشتهشد که هتروکرونی بهمنزلهی استثنائی بر قانون تکرار رئوس نیست، بلکه مهر تأییدیست بر این قانون. یکی از دانشمندانی که بر این موضوع تأکید کرد، ادوارد درینکر کوپ (Edward Drinker Cope (July 28, 1840 – April 12, 1897))، آناتومیست، خزندهشناس، ماهیشناس و دیرینهشناس مشهور قرن نوزدهم آمریکاییست.
دورهی سوم (DeBeer, 1930)
دوره استدلالی کلیدی بر ضد قانون تکرار رئوس. دو-بیر (Gavin Rylands De Beer (1899–1972)/ جانورشناس، جنینشناس و تاریخدان بریتانیایی که رابطهی میان جنینشناسی و تکامل را بررسیمیکرد. تمرکز اصلی او بر تکوین مهرهداران بهویژه جمجمهی آنها بود) مفهوم هتروکرونی را وسعت بیشتری بخشید. شواهد خلاف قانون هکل اکنون آنقدر گسترشیافتهبودند که دو-بیر آن را کلاً مردود قلمداد کند.
با این وجود وی مفهوم هتروکرونی را نگهداشت تا مجموعهای از رویدادهای متضاد با قانون تکرار رئوس توسط آن پوششدادهشود. این رویدادها تغییراتی در چارچوب زمانی تکوین برخی گونهها بودند. دو-بیر اکنون هتروکرونی را اینگونه توصیف مینمود:
بسته به تغییر یا معکوسشدن توالی مراحل تکوین، اصل هتروکرونی میتواند منجر به ظهور دیرهنگام یا زودهنگام هر ساختاری نسبت به زمانبندی سابقش طی تکوین شود. هتروکرونی هشت راه مختلف برای تغییر در نمود تکوینی ساختارها نسبت به نیاکان پیش میآورد. (DeBeer, 1930, pp. 35-36).
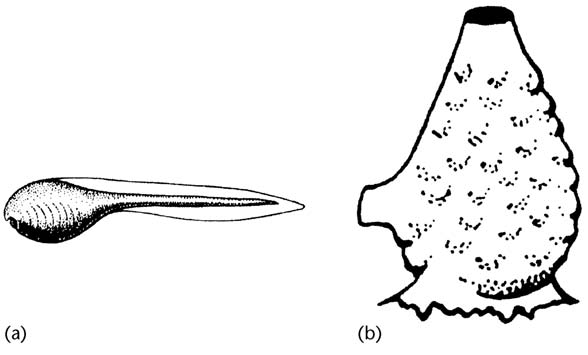
مشکل بازتعریف دو-بیر در پیچیدگی هزارتوی جدید هتروکرونی و تعاریف وابسته به آن بود. اما دو-بیر که سخنگویی کارکشتهبود از پس این پیچیدگی برآمد و برای اینکار اختصاصاً بر پدومورفوز تمرکزنمود (Cain, 2001) (Horder, 2006). درآینده توضیح بیشتری درباره پدومورفوز خواهیم داد؛ اما آنچه که در اینجا گفتنیست، مثالهاییست که میتوان بهراحتی از نمونههای پدومورفوز در طبیعت بهمیانآورد. اکسیلاتل، یکی از این مثالهاست.
مثال دیگر، تکامل مهرهداران شناگر از لاروهای قورباغهای شناگر طنابدارانیست که در دورهی بلوغ بهموجوداتی کفزی و ثابت بدلمیشوند. تکامل مهرهدارانی که تمام عمر متحرکبودند بهترین و گویاترین مثال برای نشاندادن اهمیت پدومورفوز است.
دو-بیر هشت صورت مختلف از هتروکرونی تعریفکرد، که برخی از آنها عبارتاند از سنوژنر (Caenogenesis)، نئوتنی، عقبماندگی (Retardation/ عقبماندگی، بهبیاندیگر کندشدن تکوین یکی از اندامها یا بیان ژنها) و تحلیل (Reduction/ تحلیلرفتن اندامی بر اثر کندشدن تکوین یا تسریع در توقفبیان آن). دو فرایند آخری برای توضیح اندامهای تحلیلرفته و کوچکشدن تدریجی اندامهای اضافی در طی تکامل بهکار آمدند.
نئوتنی نیز-که پیشتر توضیحدادهشد-حالتی مهم از پدومورفوز است. سنوژنز-که در زبان هکل بههر تغییر تکاملی ایجادشده نسبت به نیاکان اتلاقمیشد-اکنون به حالت خاصی از هتروکرونی گفتهمیشد که عبارتاست تغییرات بارز در ابتدای مراحل تکوین، از جمله اضافهشدن مرحلهای حدواسط چون مرحلهی لاروی که در نیاکان پیشتر وجود نداشته است.
جالباست که حتی یکی از اقسام هتروکرونی مورد نظر دو-بیر (تسریع/ Acceleration/ افزایش نرخ رشد اندامی یا بیان ژنی نسبت به بقیهی بدن)، از فرایندی منطبق با قانون تکرار رئوس هکل پیرویمیکرد. یعنی قانون هکل، اکنون صورت خاصی از هتروکرونی دو-بیر محسوب میشد.
دو-بیر بدینگونه تغییری عمیق در تفکر زیستشناسی ایجادنمود. هتروکرونی دیگر صورتی منفعل از فیلوژنی گذشتهی جانداران نبود. وی دیدگاه مکانیستیک خود را شواهد ژنتیک حمایتکرد. شواهدی که نشاندادند زمانبندی تکوین، میتواند در طی تکامل تحت تأثیر عوامل ژنتیک تغییریابد. وی بهعلاوه نشانداد هورمونهای شناختهشده در این تغییر دخیلاند (تصویر ۲).
دورهی چهارم (Gould, 1977)
آغاز دیدگاههای امروزین. زمانی که گولد در کتاب معروف خود در سال ۱۹۷۷ بهطور مشروحی بهاین موضوع پرداخت، مدتها بود که هتروکرونی-بهدلیل ابهامها و پیچیدگیهایش-از دایرهی لغات زیستشناسان خارج شده بود.
گولد، در سایهی سادگی زبانش، نشانداد چهار مورد از اقسام مورد اشارهی دو-بیر (از جمله تغییرپذیری بالغین/ Adult Variation و انحراف/ Deviation) ارتباطی به تغییر در زمانبندی تکوینی ندارند، بلکه مربوط به تمایز سلولی و تثبیت انواع جدیدی از مولکولها و بیانهای ژنی در طول تکامل هستند.
بهعلاوه وی نشانداد، در مجموع دو رویداد ظاهری بر اثر هتروکرونی محتملاست، و هر کدام از این دو، میتواند نتیجهی دو فرایند-در مجموع چهار فرایند-باشد. نخستین رویداد ظاهری مورد اشارهی وی، «تکرار رئوس» بود که یا بر اثر تسریع تکوین بدن (معادل مفهوم تسریع نزد دو-بیر)، یا عقبماندگی بلوغ جنسی (معادل مفهوم ئیپرمورفوز/ Hypermorphosis نزد دو-بیر) رخ میدهد.
اما رویداد دوم، پدومورفوز بود، که یا بر اثر بلوغ تسریعشده (که گولد آن را پروژنز/ Progensis نامید) رویمیدهد یا عقبماندگی تکوین بدن (معادل نئوتنی نزد دو-بیر). بنابراین با تحویل همهی اتفاقات حین هتروکرونی به فرایند تسریع یا عقبماندگی، گولد توانست مشروحات دو-بیر را سامانمند کند. طبق آنچه گولد بیانکرد، پروژنز متمایل به تولیدمثل زودهنگام در چرخهی زندگی و استراتژی r است؛ در مقابل ئیپرمورفوز متمایل به انتخاب K است.
بلوغ دیرهنگام در مورد دوم، میتواند با تکامل ویژگیهای بارز در جوامع پیشرفتهی مهرهداران بزرگ مرتبطباشد. گولد، بدون تغییر در بنیان تفکر دو-بیر، تنها منظر نگاه را از اندام به جاندار و از جنین به چرخهی زندگی تغییرداد. همچنین واژهی تکرار رئوس را باردیگر زندهکرد-درحالیکه دو-بیر مایلنبود از این واژه استفاده کند.
اگرچه گولد در سادهکردن مفهوم هتروکرونی کوشید، اما درعوض به فرمولبندی و رقومیسازی این مفهوم تمایلیافت. گولد، با مدل سادهی ساعت خود به این کار دستزد، اما بهزودی در مقالهای مفصلتر بهاین موضوع پرداختهشد (Alberch, et al., 1979): اکنون دورهی زندگی با کمک نمودارها و ریاضیات، بهصورت «نقشهی راه» (Trajectory) نمایشداده میشد.
مفهوم امروزین
دلیل اصلی تمایل و توجه زیاد به اهمیت تکوین در زیستشناسی تکاملی، نوشتههای گولد بود. از آن زمان تا کنون هتروکرونی مورد توجه زیادی قرارگرفتهاست؛ چرا که بیش از هر فرایند دیگری، در تکامل ریختشناخت اهمیتدارد. امروزه نگاه به هتروکرونی حتی از نگاه فرمولبندیشدهی گولد نیز فراتررفته و شاید همین نگاه گولد بود که موجب همگرایی مفهوم هتروکرونی و آلومتری (Allometry) شد. در حقیقت نرخ رشد قسمتهای مختلف بدن در دورههای مختلف در بسیاری از موجودات برابر نیست. این نابرابری در نرخ رشد قسمتهای مختلف بدن را آلومتری مینامیم.
بهترین مثال کاهش نرخ رشد اندازهی سر نسبت به نرخ رشد اندازهی بدن در طی زندگی انسان است. این تفاوت نرخ رشد، موجبمیشود انسان جوان نسبت به انسان بالغ، دارای نسبت سر به بدن بزرگتری باشد. امروزه، هتروکرونی و آلومتری دو مفهوم توأمان در نظر گرفته میشوند.
نگاهی جدیدتر به هتروکرونی مفاهیمی نو چون «هتروکرونی رشد عمومی» (Global Growth Heterochronies)، «هتروکرونی رشد موضعی» (Local Growth Heterochronies)، «هتروکرونی متوالی و سازمانی» (Sequential and Organizational Heterochronies) و «هتروکرونی تمایزی» (Differentiative Heterochrony) به زیستشناسی معرفینمود (McKinney & McNamara, 1991). مطالعات جدیدتر حتی به مراحل آغازین تکوین میپردازند و از مفاهیم سلولی و مولکولی بهره میبرند.
توصیف و تشریح
همانطور که گفتهشد، نگاه امروزین ما به هتروکرونی وامدار نگاه گولد به این موضوع است. در حقیقت، با مرور مختصری که به تاریخچهی مفاهیم هتروکرونی داشتیم، میتوان گفت بسیاری از مفاهیم امروزی را نیز مرورکردهایم. شاید بنیانیترین چیزی که نسبت به نوشتههای گولد تغییرکردهاست، اصطلاحشناسی هتروکرونی در حال حاضر است. هتروکرونی در مجموع شامل شش فرآیند کلیست، که دو نمود ظاهری ایجادمیکنند. این دو نمود عبارتند از پدومورفوز و پرامورفوز (Peramorphosis).
پدومورفوز
پدومورفوز، به ظهور و غلبهی صفاتی مشابه افراد جوانتر نیاکان گفتهمیشود. یکی از بهترین مثالهای پدومورفوز، کوتاهشدن پوزه و کوچکشدن پوزهی انسان نسبت به نیاکان پیشین خود است. اگر به روند رشد قسمتهای مختلف جمجمه در انسانریختها نظری بیافکنیم، خواهیمدید که شکل جمجمهی افراد جوانتر الگویی مانند انسانها دارد، و ویژگیهای «میمونمانند» با رشد فرد بارزتر میشود. بنابراین، تکامل انسان از چنین نیاکانی، همراهبودهاست با تثبیت ویژگیهای جمجمه و صورت افراد جوانتر نیایی، در افراد بالغ گونهی جدید. اما اینکه با چه مکانیسمی این اتفاق رخمیدهد چیزیست که در ادامه خواهیم دید.

پروژنز
در پدومورفوز سه فرایند تکاملی ممکناست باعث غلبهی صفات افراد جوان در طی تکامل شود. نخستین آنها پروژنز است. همانطور که گولد هم در این مورد بیانکردهبود، پروژنز عبارتاست از توقف زودهنگام تجلی یکی از ویژگیهایی که در فرد بالغ قرار است دیدهشود. برای توصیف دقیقتر و روشن پروژنز و دیگر فرایندهای هتروکرونیک، باید ابتدا با برخی تعاریف ساده آشنا شویم.
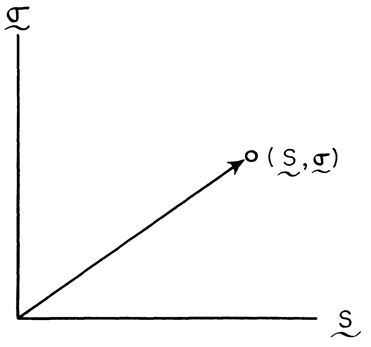
در شکل شمارهی شش، دو نمونه از رشد آلومتریک اعضای بدن آوردهشدهاست. در این شکلها به شیب متغیر رشد (شتاب متغیر نرخ رشد) توجه کنید. اگر مانند شکل شمارهی هفت، «نقشهی راه» یکی از ویژگیهای گونهی نیایی، از نقطهی α شروعشده و به نقطهی β ختمشود، شیب خط βα نشانگر رشد آلومتریک این ویژگی خواهد بود.
محورهای x و y نیز بهترتیب نمایشگر لگاریتمهای طبیعی (ln) رشد کلی بدن (ln x) و رشد ویژگی مربوطه (ln y) هستند. دلیل انتخاب لگاریتم طبیعی، خطیکردن نقشهی راه تکوین ویژگی مربوطه است-ویژگیهای آلومتریک، دارای نرخ و شتاب رشد متغیر هستند.
در صورتی که شیب خط یک باشد (با زاویهی نسبت به محور xها) بهمعنی فقدان رشد آلومتریک است و در شکل هفت که شیب خط بیش از یک است، ویژگی مربوطه (y) دارای رشد آلومتریک فزاینده نسبت به رشد کلی بدن است.


از اینجا به بعد خواهیمدید با بروز هر یک از حالات ششگانه هتروکرونیک، چه تغییراتی در نمودار نقشهی راه رشد و ظاهر حیوان فرضی ما بروز خواهد نمود.

آنگونه که در شکل شمارهی هشت دیدهمیشود، پروژنز بهسادگی توقف رشد آلومتریک ویژگیهای جانور بالغ نیاییست. نرخ رشد آلومتریک در اینجا تغییرینکرده، بلکه این محدودهی زمانی رشد است که کاسته شده است.
تأخیر
فرایند محتمل دیگر در پدومورفوز، تأخیر (Postdisplacement) در بروز صفات بلوغ است. در این حالت هم، مانند پروژنز، تغییری در نرخ رشد آلومتریک رخنمیدهد. بلکه محدودهی زمانی رشد کاستهمیشود. البته بر خلاف پروژنز که رشد آلومتریک خیلی زود متوقفمیشود، اینبار رشد دیرتر از نیاکان پیشین شروعمیشود و درنتیجه باز هم از دورهی رشد آلومتریک کاستهشده و صفت نیاکان بالغ، در گونهی جدید تحلیل خواهد رفت.

نئوتنی
نئوتنی، به دلیل تأکیدهای دو-بیر، شاید شناختهشدهترین شکل از هتروکرونی باشد. در این وضعیت، گونهی جدید نسبت به نیاکان پیشین خود، دارای سرعت رشد کمتری در مورد ویژگیهای بارز در بلوغ است. در این حالت، نرخ رشد تغییرمیکند نه مدتزمان رشد.
در هر سه حالت ذکرشده، نتیجهی کار تحلیلرفتن صفات فرد بالغ نیایی در افراد بالغ گونهی جدید است. فراموشنکنیم که در طبیعت، هیچکدام از این حالات مطلقنیستند و ممکناست ترکیبی از دو- یا سهتای آنها در تکامل گونهای تأثیرگذاشتهباشد. در حقیقت، آنچه که اغلب بهروشنی و بیگمان میتوان در موردش سخنگفت، وقوع پدومورفوز است، نه بیان دقیق اینکه کدام یک از فرایندهای سهگانهی گفتهشده منتهی به پدومورفوز شدهاند.

پرامورفوز
پرامورفوز، نمودی کاملاً متضاد با پدومورفوز دارد. بسیاری از صفات ثانویهی جنسی که نمایی اغراقشدهدارند (شاخکرگدن، گوزن، دم طاووس،…) یا صفات دیگری که نمود اغراقشدهدارند ولی لزوماً در اثر انتخاب جنسی ظاهرنشدهاند (گردن کشیدهی داینوسورهای سوروپود (Sauropod Dinosaurs/ داینوسورهایی بسیار عظیم با گردنها و دمهای کشیده)، گردن کشیدهی زرافه، خرطوم فیل،…) نیز با همین فرایند توسعه یافتهاند.
در واقع همانگونه که گولد بهدرستی در کتابش اشارهنمود، پرامورفوز در جانورانی با استراتژی K بروز مییابد. گولد برای پرامورفوز از همان عبارت قدیمیتر تکرار رئوس استفادهمیکرد. در واقع تکرار رئوس، صورت خاصی از هتروکرونیست که معنایی نزدیک به پرامورفوز مورد بحث ما دارد. پرامورفوز نیز چون پدومورفوز، بهواسطهی سه فرایند ممکناست نمودیابد. این سه فرایند عبارتند از ئیپرمورفوز، تسریع و تعجیل (Predisplacement).
ئیپرمورفوز
در ئیپرمورفوز، اتفاقی دقیقاً برعکس پروژنز میافتد. بدینمعنی که نرخرشد ثابتمیماند اما فرایند رشد طولانیتر شده و بهتدریج در گونهی جدید ویژگیهای مربوط به دورهی بلوغ، حالتی اغراقشده و بارزمییابند.

تعجیل
در مورد تعجیلنیز باید گفت با فرایندی دقیقاً دربرابر تأخیر مواجههستیم. یعنی بازهم بدون تغییر در نرخ بیان صفات، زمان شروع بیان و رشد آنها پیشافتاده و در گونههایی که تحت تأثیر این فرایند بودهاند برخی صفات ثانویهی جنسی ممکناست حتی پیش از بلوغ جنسی بروز پید اکند.

تسریع
اما مهمترین فرایندی که منتهی به بروز پرامورفوز میشود، تسریع است. دقیقاً متقابل با نئوتنی، در اینجا نرخرشد افزایشمییابد و گونهی جدید نسبت به نیاکان پیشین، صفات دورهی بلوغ را بارزتر نمایشمیدهد. همانطور که در مورد پدومورفوز نیز گفتهشده، این فرایندها بهتنهایی رخنمیدهند، بلکه معمولاست که تسریع با یکی یا هردو فرایند دیگر منتهی به پرامورفوز همراه باشد.

بنابر آنچه گفتهشد، مجموعهای از تغییر در بیان ژنها میتوانند نهایتاً به یکی از دو نمود پدومورفوز و پرامورفوز در گونههای جدیدی که تحت تأثیر این فرایندها تکاملمییابند، بروزیابد؛ و همانطور که پیشتر اشارهشد بسیاری از تغییرات مهم در جهان جانوری، بر همین اساس ظاهرشدهاند. این صفات لزوماً «اندازه»ی اندامها نیستند، بلکه ممکناست تغییر در غلظت یک هورمون یا تغییراتی ازین قسم باشند.


روندهای تکاملی
روند تکاملی، تغییری دائمی و ممتد در وضعیت یک ویژگی یا مجموعهای از ویژگیهاست که در طی زمان تکاملی، منجر به تغییراتی عظیم خواهدشد. چنین روندهایی در مقیاس گونه-بهعنوان تکامل خرد (Microevolution)-یا فراتر از آن-بهعنوان تکامل کلان-شناختهشدهاند. هتروکرونی، یکی از مهمترین عوامل ایجادکنندهی این روندهای تکاملیست.
یکی از مهمترین نمونههای حیطهی تکامل کلان، بزرگشدن اندازهی اسبها و کاهش انگشتان آنها در طی سنوزوئیک است. از نمونههای مهم دیگر بزرگشدن اندازهی بدن و مغز انسان، روند تغییر پیچیدگی خطوط درز (Suture) در آمونیتها، و حتی روند تجمیع سلولهای یوکاریوتی در جهت ایجاد کلنیها و موجودات پرسلولیست.
اغلب تغییرات تکاملی عظیم در یک روند، با بدهبستانهای ریختشناختی و عملکردی همراههستند. بسیاری از این بدهبستانها دارای مبنایی تکوینیاند (McNamara, 2006). برخی از این موارد عبارتند از افزایش اندازه و وزن بدن در پرندگان بهازای ازدستدادن قدرت پرواز، و افزایش اندازه بدن و مغز انسان در ازای پیچیدگی و اندازهی آرواره و روده. تمام این نمونهها، مثالهایی از هتروکرونیاند.
در حقیقت، هتروکرونی عامل ایجاد هر دو روند کلان و خرد تکاملیست (McNamara, 2006). از طرف دیگر، روندهای تکاملی، آوردگاه بروز هتروکرونی و تغییرات هتروکرونیک بهشمار میروند. در تکامل کلان، یعنی در مقیاس فیلوژنتیک فراتر از گونه، ممکن است با دو روند تکاملی هتروکرونیک مواجهشویم. در مورد روندهای تکاملی هتروکرونیک، بهدلیل پیچیدگی زیاد، نمیتوان قطعاً قضاوتکرد کدام یک از فرایندهای ششگانه موجب بروز روند تکاملی موردنظر شدهاست. تنها چیزی که مسلم است، تغییرات هتروکرونیک همجهت در تمامی- یا اکثر تبارهای خویشاوندیست که درگیر روند تکاملی مشترکی هستند.
دو روند هتروکرونیک، روند پدومورفیک (Paedomorphocline) و روند پرامورفیک (Peramorphocline) نامدارند (McNamara, 2006). اگربخواهیم مثال روشن و واضحی برای هرکدام بیاوریم، روند کوچکشدن اندازهی داینوسورهای گوشتخوار، در طی مزوزوئیک، تا حدی که پرندگان کوچک چند-ده گرمی از آنها تکامل یافتند، مثالی عالی برای روند پدومورفیک، و روند بزرگشدن اسبها در سنوزوئیک، مثالی عالی برای روند پرامورفیک است.
بهنظر میرسد در طبیعت، بسیاری از تغییرات تکاملی، در سایهی روندهایی فراگونهای-در مقیاس خانواده و بزرگتر-بروزمییابند و خود گونهها، تنها واحههای کوچکی در دل روندهای تکاملی بزرگ هستند که تفاوتهایی جزئی و مختصر با یکدیگر دارند.

میتوان با اطمینان گفت، روند تکاملی منجر به کوچکشدن اندازهی نیاکان شکارگر پرندگان، همیشه بر یک فرایند منطبقنبودهاست و تا پیش از تبار پرندگان، نرخ رشد اندازهی بدن دائماً افزودهشده، و این یعنی از تروپودهای ابتدایی چهار تا ۱۰ متری، تا پیش از پیدایش پرندگان، مسیری طولانی برای رسیدن به اندازهای کمتر از یک متر (روند پدومورفیک)، اما همراه با افزایش نرخ رشد (تسریع) طیشدهاست و احتمالاً دلیل بروز پدومورفوز، غلبهی اثر پدومورفیک پروژنز بر اثر پرامورفیک تسریع بوده است (تصویر ۱۷ و ۱۶).

مثال پرندگان بهدلیل وسعت و عمق تحقیقات انجامشده، شاید عینیترین و مستندترین مثالی باشد که نشانمیدهد، نمیتوان قطعاً در مورد ماهیت فرایندهای تکاملی منتهی به روند پدومورفیک یا پرامورفیک اظهارنظر کرد.
البته همهی روندهای تکاملی به همین پیچیدگی نیستند.
میتوان در برخی تبارهای بیمهرگان که ساختار بدنی سادهتری دارند، روندهای تکاملی سادهای شناساییکرد، که بهروشنی در طی فیلوژنی چند گونه و جنس، الگوی پدومورفیک یا پرامورفیک مشهود باشد.
از نمونههای معروف در مورد روند تکاملی هتروکرونیک (Heterochronocline) در بیمهرگان، میتوان به تکامل چندین ویژگی مهم در تبارهای Ammonoidea اشارهنمود. در برخی تبارها این ویژگیها (اندازهی بدن، پیچیدگی خطهای درز،…) روندی پرامورفیک و در برخی تبارهای دیگر روندی پدومورفیک نشان میدهند.
اما یکی از بهترین مثالهایی که در مورد روندهای هتروکرونیک میتوان زد، در مورد دو جنس Tegulorhynchia و Notosaria از براکیوپودهای دوران سنوزوئیک است. اهمیت این مثال در آناست که میتوان در مورد رابطهی بومشناسی و روند هتروکرونیک نظرداد. این روند احتمالاً در اثر تغییرات محیطی از آبهای عمیق به سمت آبهای کمعمق بودهاست (McNamara, 2003) (McNamara, 2006).




از موارد جالب روندهای پرامورفیک، روندهایی هستند که در اثر انتخاب جنسی ظاهرشدهاند. برای مثال، در خانوادهی Phasianidae (قرقاولها، کبکها و خروسها) انتخاب جنسی کمابیش موجب بروز صفاتی اغراقشده در رنگ پروبال و کاکلهای پری و گوشتی روی سر شدهاست. که نمونهی فوقالعادهی آن طاووس است. یکی دیگر از مثالهای مرتبط با انتخاب جنسی روند پرامورفیک توسعه و ستبر شدن شاخ در Ceratopsia است (تصویر ۲۰بـ.).

شاید یکی از بهترین نمونههای رابطهی هتروکرونی و محیط، گونههای پلانکتیک فورامینیفرها باشند. آنگونه که در تصویر ۲۰ دیدهمیشود، گونههای مناطق کمعمق که آبهایی متلاطمتر دارند، متمایل به استراتژی r و تولیدمثل زودهنگام (پدومورفیک) دارند. اما گونههای مناطق عمیقتر با چرخههای زندگی طولانیتر و پیچیدهتر، تمایل به انتخاب K دارند.

مثالهای فراوان دیگری نیز وجود دارند که نشاندهندهی رابطهی عمیق نمود هتروکرونیک در روندهای تکاملی، و تغییرات اکوسیستم، رفتار، انتخاب جنسی، و تکوین هستند. برای مثال در Ornithopoda، روند پرامورفیک به سمت بزرگشدن اندازهی بدن و آرواره و پیچیدگی فزایندهی آرواره مشهود است.
جالب است که بخشی از این این روند، مدیون تکامل همبسته (Co-evolution) با گیاهان گلداریست که در کرتاسه ظاهرشدند و موجب تکامل جداگانه و همگرای مجموعههای دندانی فشرده در آروارهی اورنیتوپودها و سراتوپسها شدند.
در برخی موارد، مانند اثر جزیرهای، تمام وقایع رخداده در تغییرات اندازهی بدن یا قسمتهای مختلف بدن، از روندی هتروکرونیک پیرویمیکند. حتی این احتمال وجود دارد برخی گروههای مهم جانوران، مانند پرندگان، در حین روند هتروکرونیک ناشی از اثر جزیرهای تکاملیافته باشند.
هرچند نکتهی مهمی در این جا وجود دارد که نباید بهسادگی از کنار آن گذشت. در تشخیص روندهای هتروکرونیک-به ویژه در مواردی مانند اثر جزیرهای که یک جنس یا حتی گونه به تنهایی تحت اثر جزیرهای تغییرمییابند-شناخت روند فیلوژنتیک مقدمهای بر تشخیص نوع اثر جزیره و روند هتروکرونیک است.
مثال بارز آن، اشتباهیست که سالها در مورد اژدهای کومودو (Varanus komodensis) رخدادهاست. غالباً گفتهشده که این سوسمار غولپیکر در اثر زندگی در جزیره، به این اندازهی بزرگدستیافته، اما نگاهی به درخت فیلوژنتیک این موجود نشانمیدهد، نزدیکترین خویشاوندان اژدهای کومودو که در سرزمین اصلی میزیستند (نه جزایر) اندازههایی نسبتاً بزرگتر داشته و دارند. بنابراین نتیجهگیریهایی که در مورد روند هتروکرونیک و پرامورفیک این گونه بودهاند، همگی اشتباههستند (Gould & McFadden, 2004).
آنچه که هتروکرونی هست، و آنچه که نیست
ممکناست بهنظر برسد نمودارها و معادلات لگاریتمی تنها جنبهی تزئینی دارند و نمیتوان از آنها استفادهی چندانی نمود. درحالیکه کاملاً برعکساست. در زیستشناسی تکاملی، مسیری روبهپیش برای ریاضیوار کردن مفاهیم وجوددارد و این مسیر، در مورد مطالعات هتروکرونیک نیز کاملاً جدی و پیگیرانه است.
در اینجا تنها برای نمونه، ابتدا چند نمودار پیشفرض (اندکی متفاوت با نمودارهایی که در تصاویر پیشین آمدهاند) ارائهمیشوند، و آنگاه با استفاده از این نمودارها، مثالی عینی از هتروکرونی، و تفاوت آن با تغییر اندازههای دیگر در طی تکامل آوردهمیشود، تا درعینحال به مفهوم ریاضی هتروکرونی نیز تأکید شده باشد.
منظور از تغییر اندازههای تکاملی دیگر بهجز هتروکرونی، تأکید بر مواردیست که قسمتهایی متفاوت از فرایند تکوینی، دچار تغییراتی متفاوت در نرخ و شدت بیان میشوند. در مقابل هتروکرونی، زمانی رخمیدهد که فرایند تکوینی ویژهای، بهطور یکدست سریعشده، کندشده، یا چهارچوب زمانبندیاش تغییر یابد.
بنابراین آنچه که ممکناست اشتباهاً هتروکرونی شناختهشود، اما هتروکرونی نیست، تغییر در سرشت فرایندهای تکوینی، به جای نرخ و چهارچوب زمانی آنهاست (Rice, 1997).

در شکل ۲۳، مفهوم ریاضی تفاوت هتروکرونی، با تغییرات دیگری که برخلاف هتروکرونی بر اساس تغییر در سرشت مسیر تکوینی ظاهرشدهاند، نشاندادهشدهاست. در نمودارهایی که پیش از این دیدهایم، اقسام هتروکرونی به کمک نمودارهای خطیشده نشاندادهشدند. در شکل ۲۴، همین اقسام را میتوان بهکمک نمودارهای غیرخطی مشاهده کرد.

اگر بخواهیم نمونهای عملی در این مورد مثالبزنیم، مقایسه نقشهی راه رشد مغز در انسان و شمپانزه و میمون مکک (Macaques (genus Macaca)/ میمونی از زیرخانوادهی Cercopithecinae، که بعد از انسان، دارای بیشترین پراکنش در راستهی نخستیها/ Primates است و ۲۲ گونه آن تاکنون شناساییشدهاند) آورده شده است (Rice, 2002).

اکنون در نمودار دیگری، مغز انسان را با میمون مکک مقایسه میکنیم.

اگر مغز شمپانزه و مکک را نیز مقایسهکنیم، همین اختلاف میان انسان و مکک، میان آنها نیز دیدهمیشود. از طرفی اگر میمون مکک را با میمون سنجابی (Squirrel Monkey (Genus Saimiri)/ از زیر خانوادهی Saimirinae از میمونهای بر جدید، ساکن آمریکای مرکزی و جنوبی) مقایسهکنیم، میبینیم که رشد مغز آنها با هم منطبق است.
بنابراین نتیجه میگیریم، پس از آخرین نیای مشترک ما، با میمونهای برقدیم (۲۵ میلیونسالپیش)، و پیش از آخرین نیای مشترک ما با شمپانزهها (هفتمیلیونسالپیش)، تغییری غیرهتروکرونیک در رشد مغز نیاکان ما رخ داده است.
اگر بخواهیم این اطلاعات را روی درخت فیلوژنتیک نشاندهیم، خواهیمدید که تغییرات اصلی و مهم مغز ما نسبت به میمونهای دمدار، پیش از اشتقاق انسانها و شمپانزهها رخدادهاند. تنها تغییر مهم بعد از جداشدن از شمپانزهها، رشد هتروکرونیک مغز انسان بوده است.

اگر بههمین ترتیب، نقشهی راه رشد وزن بدن در انسان و شمپانزه مقایسهشود، به نتایج جالبی خواهیمرسید. زیرا در نظر اول نمیتوان با هیچیک از مدلهای هتروکرونیک نقشهی راه رشد انسان را با شمپانزه انطباقداد. اما اگر رشد انسان را، پیش و پس از بلوغ، جداگانه درنظر بگیریم، هرکدام با تغییری هتروکرونیک منطبق با شمپانزه خواهد بود.

با توجه به تصویر ۲۸، میبینیم که رشد انسان نسبت به شمپانزه در کل دارای ئیپرمورفوز متوالیست (طولدورهی رشد افزودهشدهاست). بهعلاوه تا پیش از بلوغ، انسان دارای نرخ رشد کمتری نسبت به شمپانزه است (الگوی نئوتنی)؛ اما پس از بلوغ، نرخ رشد انسان، مطابق شمپانزه میشود.
این نئوتنی، همان است که پیشتر در مقایسهی شکل جمجمهی انسان و شمپانزه و مطابقت شکل جمجمهی انسان بالغ با شمپانزهی جوان متوجه آنشدهبودیم. این تغییرات هتروکرونیک باعثشده دورهی پیش از بلوغ انسان طولانیشود و زمان کافی برای آموزش نزد افراد بالغ برای کودکان فراهمگردد. بهعلاوه، در زمان بلوغ، تغییرات بسیار فاحشتر از شمپانزهها بهچشم بیآیند.
پایانبندی
بهنظر میرسد تحقیقات وسیعی در مورد هتروکرونی، بهویژه در رابطه با اکولوژی، تکامل و رفتارشناسی انجامشدهاست. اما در عوض تحقیقات تکوینی، بهعنوان رشتهی مادری که این زمینهی تحقیقاتی از دل آن زادهشد، هنوز در اول راه بررسی روندهای هتروکرونیک هستند.
هتروکرونی، نه تنها در جانوران، بلکه در گیاهان نیز دارای اهمیت بهسزاییست (Lord, 2001). در برخی گروهها مانند داینوسورها تحقیقات دامنهداری برای کشف و توصیف روندهای هتروکرونیک صورتپذیرفتهاست (Weishampel & Horner, 1994) (Long & McNamara, 1997). تکامل بسیاری از گروههای مهم جانوری اصولاً بر مبنای هتروکرونی رخدادهاست-برای مثال پرندگان (Padian, et al., 2001)، پستانداران (Luo, et al., 2001)، حشرات (Grimaldi & Engel, 2005, pp. 333-335).
گام بعدی در تحقیقات هتروکرونیک و تمامی زمینههای دیگر زیستشناسی تکاملی، بررسی مستقیم تغییرات جنینی و اثر مورفوژنها به ازای تغییرات فیلوژنتیک است. زیرا این مکانیسمهای تکوینی هستند که واسطهی تغییرات ژنوم و تغییرات آناتومیک هستند.
منابع اشاره شده در متن برای مطالعه بیشتر
- Alberch, P., Gould, S. J., Oster, G. F., & Wake, D. B. (1979). Size and shape in ontogeny and phylogeny. Paleobiology , 5 (3), 296-317.
- Cain, J. (2001). Gavin Rylands De Beer. In Encyclopedia of Life Science (pp. 1-2). John Wiley & Sons, Ltd. www.els.net.
- Caron, M., & Homewood, P. (1983). Evolution of Early Planktic Foraminifers. Marine Micropalaeontology , 7, 453-462.
- DeBeer, G. R. (1930). Embryology and Evolution. Oxford: Clarendon Press.
- Gould, G. C., & McFadden, B. J. (2004). Gigantism, Dwarfism, and Cope's Rule: Nothing in Evolution Makes Sense Without a Phylogeny. Bulletin of American Museum of Natural History , 285, 219-237.
- Gould, S. J. (1977). Ontogeny and Phylogeny. Cambridge, MA: Harvard University Press.
- Grimaldi, D., & Engel, M. S. (2005). Evolution of the Insects. New York: Cambridge University Press.
- Horder, T. (2006). Heterochrony. In Encyclopedia of Life Science (pp. 1-6). John Wiley & Sons, Ltd. www.els.net.
- Long, J. A., & McNamara, K. J. (1997). Heterochrony. In P. J. Currie, & K. Padian (Eds.), Encyclopedia of Dinosaurs (pp. 311-317). London: Acaedemic Press.
- Lord, E. M. (2001). Heterochrony in Plants. In Encyclopedia of Life Science (pp. 1-4). John Wiley & Sons, Ltd. www.els.net.
- Luo, Z. X., Crompton, A. W., & Sun, A. L. (2001). A New Mammaliaform from the Early Jurassic and Evolution of Mammalian Characteristics. Science , 292, 1535-1540.
- McKinney, M. L., & McNamara, K. J. (1991). Heterochrony: The Evolution of Ontogeny. New York: Plenum Press.
- McNamara, K. J. (1990). Heterochrony. In D. E. Briggs, & P. R. Crowther, Palaeobiology (pp. 111-119). Oxford: Blackwell Science Ltd.
- McNamara, K. J. (2003). Iportance of Heterochrony. In D. E. Briggs, & P. R. Crowther, Palaeobiology II (pp. 180-188). Oxford: Blackwell Science Ltd.
- McNamara, K. J. (2006). Evolutionary Trends. In Encyclopedia of Life Science (pp. 1-7). John Wiley & Sons, Ltd. www.els.net.
- Padian, K., De Ricqles, A. K., & Horner, J. R. (2001). Dinosaurian growth rates and bird origins. Nature , 412, 405-408.
- Rice, S. H. (1997). The Analysis of Ontogenetic Trajectories: When a Change in Size or Shape Is Not Heterochrony. PNAS , 94, 907-912.
- Rice, S. H. (2002). The Role of Heterochrony in Primate Brain Evolution. In N. Minugh-Purvis, & K. J. McNamara (Eds.), Human Evolution Through Developmental Change. Baltimore: Johns Hopkins University Press.
- Sereno, P. C. (1997). The Origin and Evolution of Dinosaurs. Annual Review of Earth and Planetary Science , 25, 435-489.
- Weishampel, D. B., & Horner, J. R. (1994). Life History Syndromes, Heterochrony, and the Evolution of Dinosauria. In K. Carpenter, K. F. Hirsch, & J. R. Horner (Eds.), Dinosaur Eggs and Babies (pp. 229-243). New York: Cambridge University Press.
اگر این مطلب برای شما مفید بوده است، آموزشهای زیر نیز به شما پیشنهاد میشوند:










سلام استاد خسته نباشید و خداقوت، ببخشید میشه لطفا یه دوره مقاله در مورد تحلیل گروه های متنوع بی مهره ها و ویژگی هایشان و رده ی پستانداران و راسته ی پریمات ها و تکامل انسان و هومونیده برایمان بنویسین لطفا اگر صلاح میدونین؟ خیلی ممنون میشوم زیرا مقالاتتان واقعا عالی ودرجه یک و بسیار علمی هستند. خیلی ممنون